
Limbo
Descent of Christ to Limbo - Сошествие Христа во ад.
Слово "limbo". Англо-русский словарь Мюллера.
1. религиозное — лимб, преддверие Ада.
2. заточение, тюрьма.
“No,” said the other priest; “reason is always reasonable, even in the last limbo, in the lost borderland of things.
— Нет, — сказал отец Браун, — разум разумен везде.
Сапфировый крест. Честертон Гилберт Кийт, стр. 17
3. склад ненужных вещей.
4. пребывание в забвении.
Millions of refugees are living in exile, in limbo.
Они живут в чужих странах, спасаясь от войны,
Субтитры видеоролика "Судно с 500 беженцами затонуло в море. История двух выживших. Melissa Fleming", стр. 3
Лимбическая система
Лимбическая система (от лат. limbus — граница, край) — совокупность ряда структур головного мозга. Окутывает верхнюю часть ствола головного мозга, будто поясом, и образует его край (лимб). Участвует в регуляции функций внутренних органов, обоняния, автоматической регуляции, эмоций, памяти, сна, бодрствования и др.[источник не указан 2292 дня] Термин лимбическая система впервые введён в научный оборот в 1952 году американским исследователем Паулем Мак-Лином.[1]
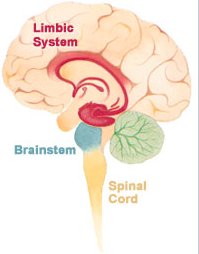
Включает в себя:
- обонятельную луковицу (Bulbus olfactorius)
- обонятельный тракт (Tractus olfactorius)
- обонятельный треугольник (Trigonum olfactorium)
- переднее продырявленное вещество (Substantia perforata anterior)
- поясная извилина (Gyrus Cinguli) (англ. Cingulate gyrus): автономные функции регуляции частоты сердцебиений и кровяного давления;
- парагиппокампальная извилина (Gyrus parahippocampalis)
- зубчатая извилина (Gyrus dentatus)
- гиппокамп (Hippocampus): требуемый для формирования долговременной памяти, обработки и хранения пространственной информации
- миндалевидное тело (Corpus amygdaloideum) (англ. Amygdala): агрессия и осторожность, страх
- гипоталамус (Hypothalamus): регулирует автономную нервную систему через гормоны, голод, жажду, половое влечение, цикл сна и пробуждения
- сосцевидное тело (Corpus Mamillare) (англ. Mammilary body): важен для формирования памяти
- ретикулярную формацию среднего мозга (Formatio reticularis)
Функции лимбической системы
Получая информацию о внешней и внутренней средах организма, лимбическая система запускает вегетативные и соматические реакции, обеспечивающие адекватное приспособление организма к внешней среде и сохранение гомеостаза. Частные функции лимбической системы:
- регуляция функции внутренних органов (через гипоталамус);
- формирование мотиваций, эмоций, поведенческих реакций;
- играет важную роль в обучении;
- обонятельная функция;
- организация кратковременной и долговременной памяти, в том числе пространственной;
- участие в формировании ориентировочно-исследовательской деятельности (синдром Клювера — Бюси);
- организация простейшей мотивационно-информационной коммуникации (речи);
- участие в механизмах сна.
------
1 «Психофизиология» / под ред. Александрова Ю. И. — 3-е издание, СПб.: «Питер», 2011 — стр. 150. ISBN 5-94723-732-6
* * *
Лимбическая система человека имеет замкнутую структуру, основанную на восходящих и нисходящих путях. Особенности ее строения заключаются в стабильных нейронных связях, которые поддерживают ее функционирование, обеспечивают продолжительное поддержание нервного возбуждения в клетках. Благодаря этому поддерживается замкнутый круг функционирования ее структур.
* * *
Лимбическая система отвечает за интеллектуальную и эмоциональную сферы деятельности человек.
* * *
Глава 4. НЕРВНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Лимбическая система
Лимбическая система представляет собой функциональное объединение структур мозга, участвующих в организации эмоционально-мотивационного поведения, таких как пищевой, половой, оборонительный инстинкты. Эта система участвует в организации цикла бодрствование—сон.
Лимбическая система как филогенетически древнее образование оказывает регулирующее влияние на кору большого мозга и подкорковые структуры, устанавливая необходимое соответствие уровней их активности.
Морфофункциональная организация. Структуры лимбической системы включают в себя 3 комплекса. Первый комплекс — древняя кора (препериформная, периамигдалярная, диагональная кора), обонятельные луковицы, обонятельный бугорок, прозрачная перегородка (рис. 4.12).
Вторым комплексом структур лимбической системы является старая кора, куда входят гиппокамп, зубчатая фасция, поясная извилина.
Третий комплекс лимбической системы — структуры островковой коры, парагиппокамповая извилина.
И, наконец, в лимбическую систему включают подкорковые структуры: миндалевидные тела, ядра прозрачной перегородки, переднее таламическое ядро, сосцевидные тела.
Особенностью лимбической системы является то, что между ее структурами имеются простые двусторонние связи и сложные пути, образующие множество замкнутых кругов (см. рис. 4.12). Такая организация создает условия для длительного циркулирования одного и того же возбуждения в системе и тем самым для сохранения в ней единого состояния и навязывание этого состояния другим системам мозга.
В настоящее время хорошо известны связи между структурами мозга, организующие круги, имеющие свою функциональную специфику. К ним относится круг Пейпеса (гиппокамп à сосцевидные тела à передние ядра таламуса à кора поясной извилины à парагиппокампова извилина à гиппокамп). Этот круг имеет отношение к памяти и процессам обучения.
Другой круг (миндалевидное тело à гипоталамус à мезенцефальные структуры à миндалевидное тело) регулирует агрессивно-оборонительные, пищевые и сексуальные формы поведения.
Считается, что образная (иконическая) память формируется кортико-лимбико-таламо-кортикальным кругом. Круги разного функционального назначения связывают лимбическую систему со многими структурами центральной нервной системы, что позволяет последней реализовать функции, специфика которых определяется включенной дополнительной структурой.
Например, включение хвостатого ядра в один из кругов лимбической системы определяет ее участие в организации тормозных процессов высшей нервной деятельности.
Большое количество связей в лимбической системе, своеобразное круговое взаимодействие ее структур создают благоприятные условия для реверберации возбуждения по коротким и длинным кругам. Это, с одной стороны, обеспечивает функциональное взаимодействие частей лимбической системы, с другой — создает условия для запоминания. Обилие связей лимбической системы со структурами центральной нервной системы затрудняет выделение функций мозга, в которых она не принимала бы участия. Так, лимбическая система имеет отношение к регулированию уровня реакции автономной, соматической систем при эмоционально-мотивационной деятельности, регулированию уровня внимания, восприятия, воспроизведения эмоционально значимой информации. Лимбическая система определяет выбор и реализацию адаптационных форм поведения, динамику врожденных форм поведения, поддержание гомеостаза, генеративных процессов. Наконец, она обеспечивает создание эмоционального фона, формирование и реализацию процессов высшей нервной деятельности.
Нужно отметить, что древняя и старая кора лимбической системы имеет прямое отношение к обонятельной функции. В свою очередь обонятельный анализатор, как самый древний из анализаторов, является неспецифическим активатором всех видов деятельности коры большого мозга.
Некоторые авторы называют лимбическую систему висцеральным мозгом, т. е. структурой ЦНС, участвующей в регуляции деятельности внутренних органов. И действительно, миндалевидные тела, прозрачная перегородка, обонятельный мозг при их возбуждении изменяют активность вегетативных систем организма в соответствии с условиями окружающей среды. Это стало возможно благодаря установлению морфологических и функциональных связей с более молодыми образованиями мозга, обеспечивающими взаимодействие экстероцептивных, интероцептивных систем и коры височной доли.
Наиболее полифункциональными образованиями лимбической системы являются гиппокамп и миндалевидные тела. Физиология этих структур наиболее изучена.
Глава 3. ПРИНЦИПЫ ОРГАНИЗАЦИИ УПРАВЛЕНИЯ ФУНКЦИЯМИ
УПРАВЛЕНИЕ В ЖИВЫХ ОРГАНИЗМАХ
Организм как единое целое может существовать только при условии, когда составляющие его органы и ткани функционируют с такой интенсивностью и в таком объеме, которые обеспечивают адекватное уравновешивание со средой обитания. По словам И. П. Павлова, живой организм — сложная обособленная система, внутренние силы которой постоянно уравновешиваются с внешними силами окружающей среды. В основе уравновешивания лежат процессы регуляции, управления физиологическими функциями.
Управление, или регуляция, в живых организмах представляет собой совокупность процессов, обеспечивающих необходимые режимы функционирования, достижение определенных целей или полезных для организма приспособительных результатов. Управление возможно при наличии взаимосвязи органов и систем организма. Процессы регуляции охватывают все уровни организации системы: молекулярный, субклеточный, клеточный, органный, системный, организменный, надорганизменный (популяционный, экосистемный, биосферный). Законы управления в сложных системах изучает кибернетика — наука об общих принципах управления в машинах, живых системах и обществе. Медицинская, физиологическая кибернетика изучает процессы управления в живых организмах.
Принципы управления. С позиций медицинской кибернетики, управление в живых организмах осуществляется управляющей системой. Она включает в себя датчики, воспринимающие информацию на входе (сенсорные рецепторы) и выходе (рецепторы исполнительных структур) системы, входные и выходные каналы связи (жидкие среды организма, нервные проводники), управляющее устройство (центральная нервная система), частью которого является запоминающее устройство (аппараты памяти). Информация, фиксированная в аппаратах памяти, определяет «настройку» системы управления на переработку определенных сведений, поставляемых через каналы связи.
Управление осуществляется с использованием двух основных принципов: 1) по рассогласованию (отклонению); 2) по возмущению.
Управление по рассогласованию предусматривает наличие механизмов, способных определить разность между задаваемым и фактическим значением регулируемой величины или функции. Эта разность используется для выработки регулирующего воздействия на объект регуляции, которое уменьшает величину отклонения. Примером такого управления является стимуляция образования глюкозы при уменьшении ее содержания в крови. Это уменьшение определяется клетками гипоталамуса, которые стимулируют выработку адренокортикотропного гормона в гипофизе. Последний усиливает образование глюкокортикоидов (кортизола) в надпочечниках. Кортизол стимулирует в печени образование глюкозы из аминокислот (глюконеогенез), что приводит к восстановлению нормального содержания глюкозы в плазме крови.
Управление по возмущению предусматривает использование самого возмущения для выработки, компенсирующего воздействия, в результате которого регулируемый показатель возвращается к исходному состоянию. Например, уменьшение парциального давления О2 в атмосферном воздухе при подъеме на высоту является возмущающим воздействием для системы дыхания, обеспечивающей оптимальное для метаболизма содержание кислорода в крови. Увеличение частоты и глубины дыхания, скорости кровотока, количества эритроцитов в крови отражает процессы регуляции по возмущению, направленные на восстановление исходных показателей содержания кислорода.
Способы управления в организме. Основные способы управления в живом организме предусматривают запуск (инициацию), коррекцию и координацию физиологических процессов.
Запуск представляет собой процесс управления, вызывающий переход функции органа от состояния относительного покоя к деятельному состоянию или от активной деятельности к состоянию покоя. Например, при определенных условиях центральная нервная система инициирует работу пищеварительных желез, фазные сокращения скелетной мускулатуры, процессы мочевыведения, дефекации и др.
Коррекция позволяет управлять деятельностью органа, осуществляющего физиологическую функцию в автоматическом режиме или инициированную поступлением управляющих сигналов. Примером может служить коррекция работы сердца центральной нервной системой посредством влияний, передаваемых по блуждающим и симпатическим нервам.
Координация предусматривает согласование работы нескольких органов или систем одновременно для получения полезного приспособительного результата. Например, для осуществления акта прямохождения необходима координация работы мышц и центров, обеспечивающих перемещение нижних конечностей в пространстве, смещение центра тяжести тела, изменение тонуса скелетных мышц.
Механизмы управления. Условно можно разделить на гуморальный и нервный.
Гуморальный механизм управления предусматривает изменение физиологической активности органов и систем под влиянием химических веществ, доставляемых через жидкие среды организма (интерстициальная жидкость, лимфа, кровь, цереброспинальная жидкость и др.). Гуморальный механизм управления является древнейшей формой взаимодействия клеток, органов и систем, поэтому в организме человека и высших животных можно найти различные варианты гуморального механизма регуляции, отражающие в известной мере его эволюцию. Одним из простейших вариантов является изменение деятельности клеток под влиянием продуктов обмена веществ. Последние могут изменять работу клетки, из которой происходит выделение этих продуктов, и других органов, расположенных на достаточном удалении. Например, под влиянием СО2, образующегося в тканях в результате утилизации кислорода, изменяется активность центра дыхания и как следствие — глубина и частота дыхания. Под влиянием адреналина, выделяемого в кровь из надпочечников, изменяются частота и сила сердечных сокращений, тонус периферических сосудов, ряд функций центральной нервной системы, интенсивность обменных процессов в скелетных мышцах, увеличиваются коагуляционные свойства крови.
Для гуморального механизма управления характерны относительно медленное распространение и диффузный характер управляющих воздействий, низкая надежность осуществления связи.
Нервный механизм управления предусматривает изменение физиологических функций под влиянием управляющих воздействий, передаваемых из центральной нервной системы по нервным волокнам к органам и системам организма. Нервный механизм является более поздним продуктом эволюции по сравнению с гуморальным, он более сложен и более совершенен. Для него характерна высокая скорость распространения и точная передача объекту регулирования управляющих воздействий, высокая надежность осуществления связи.
В естественных условиях нервный и гуморальный механизмы работают как единый нейрогуморальный механизм управления.
Нейрогуморальный механизм управления представляет собой комбинированную форму, в которой одновременно используются гуморальный и нервный механизмы; оба взаимосвязаны и взаимообусловлены. Так, передача управляющих воздействий с нерва на иннервируемые структуры осуществляется с помощью химических посредников — медиаторов, действующих на специфические рецепторы. Еще более тесная и сложная связь обнаружена в некоторых ядрах гипоталамуса. Нервные клетки этих ядер приходят в активное состояние при изменении химических и физико-химических показателей крови. Активность этих клеток вызывает образование и выделение химических факторов, стимулирующих восстановление исходных характеристик крови. Так, на повышение осмотического давления плазмы крови реагируют специальные нервные клетки супраоптического ядра гипоталамуса, активность которых приводит к выделению в кровь антидиуретического гормона, усиливающего реабсорбцию воды в почках, что обусловливает снижение осмотического давления.
Взаимодействие гуморального и нервного механизмов создает интегративный вариант управления, способный обеспечить адекватное изменение функций от клеточного до организменного уровней при изменении внешней и внутренней среды.
Средства управления. Управление физиологическими функциями осуществляется посредством передачи информации. Информация может содержать сообщение о наличии возмущающих воздействий, отклонение функций. Она передается по афферентным (чувствительным) каналам связи. Информация, передаваемая по эфферентным (исполнительным) каналам связи, содержит сообщение о том, какие функции и в каком направлении следует изменять.
Гуморальный механизм в качестве средств управления и передачи информации использует химические вещества — продукты обмена веществ, простагландины, регуляторные пептиды, гормоны и др. Так, накопление молочной кислоты в мышцах при физической нагрузке является источником информации о недостатке кислорода.
Нервный механизм в качестве средства управления, передачи информации использует потенциалы возбуждения (ПД, импульсы), которые объединяются в определенные паттерны («рисунки» возбуждения) по частоте, набору в «пачках», характеристикам межимпульсных интервалов и кодируют необходимую информацию. Показано, что паттерны возбуждений гипоталамических нейронов при формировании мотивации голода специфичны и существенно отличаются от столь же специфичных паттернов возбуждений нейронов, ответственных за формирование мотивации жажды.
Формы управления. Гуморальный и нервный механизмы предусматривают использование нескольких форм управления. Аутокринная, паракринная и гуморальная формы характерны для эволюционно более древнего механизма.
Аутокринная форма управления предполагает изменение функции клетки химическими субстратами, выделяемыми в межклеточную среду самой клеткой.
Паракринная форма управления основана на выделении клетками химических средств управления в межтканевую жидкость. Химические субстраты, распространяясь по межтканевым пространствам, могут управлять функцией клеток, расположенных на некотором удалении от источника управляющих воздействий.
Гуморальная форма управления реализуется при выделении биологических веществ в кровь. С током крови эти вещества достигают всех органов и тканей.
В основе нервного механизма управления лежит рефлекс — ответная реакция организма на изменения внутренней и внешней среды, осуществляемая при участии центральной нервной системы. Управление посредством рефлексов предусматривает использование двух форм.
Местные рефлексы осуществляются через ганглии автономной нервной системы, которые рассматриваются как нервные центры, вынесенные на периферию. За счет местных рефлексов происходит управление, например моторной и секреторной функциями тонкой и толстой кишки.
Центральные рефлексы протекают с обязательным вовлечением различных уровней центральной нервной системы (от спинного мозга до коры большого мозга). Примером таких рефлексов является выделение слюны при раздражении рецепторов полости рта, опускание века при раздражении склеры глаза, отдергивание руки при раздражении кожи пальцев и др.
В естественных условиях нервный и гуморальный механизмы едины и, образуя нейрогуморальный механизм, реализуются в разнообразных комбинациях, наиболее полно обеспечивающих адекватное уравновешивание организма со средой обитания. Например, физиологически активные вещества, поступая в кровь, несут информацию в ЦНС об отклонении какой-либо функции. Под влиянием этой информации формируется поток управляющих нервных импульсов к эффекторам для коррекции отклонения.
В других случаях поступление информации в ЦНС по нервным каналам приводит к выделению гормонов, корригирующих возникшие отклонения. Нейрогуморальный механизм создает в процессах управления многозвенные кольцевые связи, где различные формы гуморального механизма сменяются и дополняются нервными, а последние обеспечивают включение гуморальных.
Глава 3. ПРИНЦИПЫ ОРГАНИЗАЦИИ УПРАВЛЕНИЯ ФУНКЦИЯМИ
САМОРЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
В процессе эволюции живых организмов внутренняя среда была отделена от внешней и приобрела устойчивый, консервативный характер.
Французский исследователь К. Бернар писал, что условием свободного поведения живого организма является постоянство внутренней среды. По его мнению, все жизненные процессы имеют одну цель — поддержание постоянства условий жизни во внутренней среде организма. Позднее эта мысль нашла воплощение в трудах американского физиолога У. Кеннона в форме учения о гомеостазе.
Гомеостаз — относительное динамическое постоянство внутренней среды и устойчивость физиологических функций организма. Основным механизмом поддержания гомеостаза является саморегуляция.
Саморегуляция представляет собой такой вариант управления, при котором отклонение какой-либо физиологической функции или характеристик (констант) внутренней среды от уровня, обеспечивающего нормальную жизнедеятельность, является причиной возвращения этой функции (константы) к исходному уровню. В ходе естественного отбора живыми организмами выработаны общие механизмы управления процессами приспособления к среде обитания различной физиологической природы (эндокринные, нейрогуморальные, иммунологические и др.), направленные на обеспечение относительного постоянства внутренней среды. У человека и высших животных гомеостатические механизмы достигли совершенства.
Практически все характеристики внутренней среды (константы) организма непрерывно колеблются относительно средних уровней, оптимальных для протекания устойчивого обмена веществ. Эти уровни отражают потребность клеток в необходимом количестве исходных продуктов обмена. Допустимый диапазон колебаний для разных констант различен. Незначительные отклонения одних констант могут приводить к существенным нарушениям обменных процессов — это так называемые жесткие константы. К ним относятся, например, осмотическое давление, величина водородного показателя (рН), содержание глюкозы, О2, СО2 в крови.
Другие константы могут варьировать в довольно широком диапазоне без существенных нарушений физиологических функций — это так называемые пластичные константы. К их числу относят количество и соотношение форменных элементов крови, объем циркулирующей крови, скорость оседания эритроцитов.
Процессы саморегуляции основаны на использовании прямых и обратных связей. Прямая связь предусматривает выработку управляющих воздействий на основании информации об отклонении константы или действии возмущающих факторов. Например, раздражение холодным воздухом терморецепторов кожи приводит к увеличению процессов теплопродукции.
Обратные связи заключаются в том, что выходной, регулируемый сигнал о состоянии объекта управления (константы или функции) передается на вход системы. Различают положительные и отрицательные обратные связи. Положительная обратная связь усиливает управляющее воздействие, позволяет управлять значительными потоками энергии, потребляя незначительные энергетические ресурсы. Примером может служить увеличение скорости образования тромбина при появлении некоторого его количества на начальных этапах коагуляционного гемостаза.
Отрицательная обратная связь ослабляет управляющее воздействие, уменьшает влияние возмущающих факторов на работу управляющих объектов, способствует возвращению измененного показателя к стационарному уровню. Например, информация о степени натяжения сухожилия скелетной мышцы, поступающая в центр управления функций этой мышцы от рецепторов Гольджи, ослабляет степень возбуждения центра, чем предохраняет мышцу от развития избыточной силы сокращения. Отрицательные обратные связи повышают устойчивость биологической системы — способность возвращаться к первоначальному состоянию после прекращения возмущающего воздействия.
В организме обратные связи построены по принципу иерархии (подчиненности) и дублирования. Например, саморегуляция работы сердечной мышцы предусматривает наличие обратных связей от рецепторов самой сердечной мышцы, рецепторных полей магистральных сосудов, рецепторов, контролирующих уровень тканевого дыхания, и др.
Гомеостаз организма в целом обеспечивается согласованной содружественной работой различных органов и систем, функции которых поддерживаются на относительно постоянном уровне процессами саморегуляции.
Глава 3. ПРИНЦИПЫ ОРГАНИЗАЦИИ УПРАВЛЕНИЯ ФУНКЦИЯМИ
СИСТЕМНАЯ ОРГАНИЗАЦИЯ УПРАВЛЕНИЯ. ФУНКЦИОНАЛЬНЫЕ СИСТЕМЫ И ИХ ВЗАИМОДЕЙСТВИЕ
Представление о саморегуляции физиологических функций нашло наиболее полное отражение в теории функциональных систем, разработанной академиком П. К. Анохиным. Согласно этой теории, уравновешивание организма со средой обитания осуществляется самоорганизующимися функциональными системами.
Функциональные системы (ФС) представляют собой динамически складывающийся саморегулирующийся комплекс центральных и периферических образований, обеспечивающий достижение полезных приспособительных результатов.
Результат действия любой ФС представляет собой жизненно важный адаптивный показатель, необходимый для нормального функционирования организма в биологическом и социальном плане. Отсюда вытекает системообразующая роль результата действия. Именно для достижения определенного адаптивного результата складываются ФС, сложность организации которых определяется характером этого результата.
Многообразие полезных для организма приспособительных результатов может быть сведено к нескольким группам: 1) метаболические результаты, являющиеся следствием обменных процессов на молекулярном (биохимическом) уровне, создающими необходимые для жизнедеятельности субстраты или конечные продукты; 2) гомеопатические результаты, представляющие собой ведущие показатели жидких сред организма: крови, лимфы, интерстициальной жидкости (осмотическое давление, рН, содержание питательных веществ, кислорода, гормонов и т. д.), обеспечивающие различные стороны нормального обмена веществ; 3) результаты поведенческой деятельности животных и человека, удовлетворяющие основные метаболические, биологические потребности: пищевые, питьевые, половые и др.; 4) результаты социальной деятельности человека, удовлетворяющие социальные (создание общественного продукта труда, охрана окружающей среды, защита отечества, обустройство быта) и духовные (приобретение знаний, творчество) потребности.
В состав каждой ФС включаются различные органы и ткани. Объединение последних в ФС осуществляется результатом, ради достижения которого создается ФС. Этот принцип организации ФС получил название принципа избирательной мобилизации деятельности органов и тканей в целостную систему. Например, для обеспечения оптимального для метаболизма газового состава крови происходит избирательная мобилизация в ФС дыхания деятельности легких, сердца, сосудов, почек, кроветворных органов, крови.
Включение отдельных органов и тканей в ФС осуществляется по принципу взаимодействия, который предусматривает активное участие каждого элемента системы в достижении полезного приспособительного результата.
В приведенном примере каждый элемент активно способствует поддержанию газового состава крови: легкие обеспечивают газообмен, кровь связывает и транспортирует О2 и СО2, сердце и сосуды обеспечивают необходимую скорость движения крови и величину.
Для достижения результатов различного уровня формируются и разноуровневые ФС. ФС любого уровня организации имеет принципиально однотипную структуру, которая включает в себя 5 основных компонентов: 1) полезный приспособительный результат; 2) акцепторы результата (аппараты контроля); 3) обратную афферентацию, поставляющую информацию от рецепторов в центральное звено ФС; 4) центральную архитектонику — избирательное объединение нервных элементов различных уровней в специальные узловые механизмы (аппараты управления); 5) исполнительные компоненты (аппараты реакции) — соматические, вегетативные, эндокринные, поведенческие. Схема функциональной системы по П. К. Анохину представлена на рис. 3.1.
Состояние внутренней среды постоянно контролируется соответствующими рецепторами. Источником изменения параметров внутренней среды организма является непрерывно текущий в клетках процесс обмена веществ (метаболизм), сопровождающийся потреблением исходных и образованием конечных продуктов. Любое отклонение параметров от показателей, оптимальных для метаболизма, равно как и изменение результатов иного уровня, воспринимается рецепторами. От последних информация передается звеном обратной связи в соответствующие нервные центры. На основе поступающей информации происходит избирательное вовлечение в данную ФС структур различных уровней центральной нервной системы для мобилизации исполнительных органов и систем (аппаратов реакции). Деятельность последних приводит к восстановлению необходимого для метаболизма или социальной адаптации результата.
Организация различных ФС в организме принципиально одинакова. В этом заключается принцип изоморфизма ФС.
Вместе с тем в их организации есть и отличия, которые обусловлены характером результата. ФС, определяющие различные показатели внутренней среды организма, генетически детерминированы, часто включают в себя только внутренние (вегетативные, гуморальные) механизмы саморегуляции. К их числу можно отнести ФС, определяющие оптимальный для метаболизма тканей уровень массы крови, форменных элементов, реакции среды (рН), кровяного давления. Другие ФС гомеостатического уровня включают в себя и внешнее звено саморегуляции, предусматривающее взаимодействие организма с внешней средой. В работе некоторых ФС внешнее звено играет относительно пассивную роль источника необходимых субстратов (например, кислорода для ФС дыхания), в других внешнее звено саморегуляции активно и включает целенаправленное поведение человека в среде обитания, направленное на ее преобразование. К их числу относится ФС, обеспечивающая оптимальный для организма уровень питательных веществ, осмотического давления, температуры тела.
ФС поведенческого и социального уровня чрезвычайно динамичны по своей организации и формируются по мере возникновения соответствующих потребностей. В таких ФС внешнее звено саморегуляции играет ведущую роль. Вместе с тем поведение человека определяется и корригируется генетически, индивидуально приобретенным опытом, а также многочисленными возмущающими воздействиями. Примером таких ФС является производственная деятельность человека по достижению социально значимого для общества и индивида результата: творчество ученых, художников, писателей.
Аппараты управления ФС. По принципу изоморфизма построена и центральная архитектоника (аппараты управления) ФС, складывающаяся из нескольких стадий (см. рис. 3.1). Исходной является стадия афферентного синтеза. В ее основе лежит доминирующая мотивация, возникающая на базе наиболее значимой в данный момент потребности организма. Возбуждение, создаваемое доминирующей мотивацией, мобилизует генетический и индивидуально приобретенный опыт (память) по удовлетворению данной потребности. Информация о состоянии среды обитания, поставляемая обстановочной афферентацией, позволяет в конкретной обстановке оценить возможность и при необходимости скорректировать прошлый опыт удовлетворения потребности. Взаимодействие возбуждений, создаваемых доминирующей мотивацией, механизмами памяти и обстановочной афферентацией, создает состояние готовности (предпусковой интеграции), необходимое для получения адаптивного результата. Пусковая афферентация переводит систему из состояния готовности в состояние деятельности. В стадии афферентного синтеза доминирующая мотивация определяет, что делать, память — как делать, обстановочная и пусковая афферентация — когда делать, чтобы достичь необходимого результата.
Стадия афферентного синтеза завершается принятием решения. В этой стадии из многих возможных избирается единственный путь для удовлетворения ведущей потребности организма. Происходит ограничение степеней свободы деятельности ФС.
Вслед за принятием решения формируются акцептор результата действия и программа действия. В акцепторе результатов действия программируются все основные черты будущего результата действия. Это программирование происходит на основе доминирующей мотивации, которая извлекает из механизмов памяти необходимую информацию о характеристиках результата и путях его достижения. Таким образом, акцептор результатов действия представляет собой аппарат предвидения, прогнозирования, моделирования итогов деятельности ФС, где моделируются и сопоставляются параметры результата с афферентной моделью. Информация о параметрах результата поставляется с помощью обратной афферентации.
Программа действия (эфферентный синтез) представляет собой согласованное взаимодействие соматических, вегетативных и гуморальных компонентов в целях успешного достижения полезного приспособительного результата. Программа действия формирует необходимый приспособительный акт в виде определенного комплекса возбуждений в ЦНС до начала его реализации в виде конкретных действий. Эта программа определяет включение эфферентных структур, необходимых для получения полезного результата.
Необходимое звено в работе ФС — обратная афферентация. С ее помощью оцениваются отдельные этапы и конечный результат деятельности систем. Информация от рецепторов поступает по афферентным нервам и гуморальным каналам связи к структурам, составляющим акцептор результата действия. Совпадение параметров реального результата и свойств заготовленной в акцепторе его модели означает удовлетворение исходной потребности организма. Деятельность ФС на этом заканчивается. Ее компоненты могут быть использованы в других ФС. При несовпадении параметров результата и свойств модели, заготовленной на основании афферентного синтеза в акцепторе результатов действия, возникает ориентировочно-исследовательская реакция. Она приводит к перестройке афферентного синтеза, принятию нового решения, уточнению характеристик модели в акцепторе результатов действия и программы по их достижению. Деятельность ФС осуществляется в новом, необходимом для удовлетворения ведущей потребности направлении.
Принципы взаимодействия ФС. В организме работает одновременно несколько функциональных систем, что предусматривает их взаимодействие, которое строится на определенных принципах.
Принцип системогенеза предполагает избирательное созревание и инволюцию функциональных систем. Так, ФС кровообращения, дыхания, питания и их отдельные компоненты в процессе онтогенеза созревают и развиваются раньше других ФС.
Принцип мультипараметрического (многосвязного) взаимодействия определяет обобщенную деятельность различных ФС, направленную на достижение многокомпонентного результата. Например, параметры гомеостаза (осмотическое давление, КОС и др.) обеспечиваются самостоятельными ФС, которые объединяются в единую обобщенную ФС гомеостаза. Она и определяет единство внутренней среды организма, а также ее изменения вследствие процессов обмена веществ и активной деятельности организма во внешней среде. При этом отклонение одного показателя внутренней среды вызывает перераспределение в определенных соотношениях других параметров результата обобщенной ФС гомеостаза.
Принцип иерархии предполагает, что ФС организма выстраиваются в определенный ряд в соответствии с биологической или социальной значимостью. Например, в биологическом плане доминирующее положение занимает ФС, обеспечивающая сохранение целостности тканей, затем — ФС питания, воспроизведения и др. Деятельность организма в каждый временной период определяется доминирующей ФС в плане выживания или адаптации организма к условиям существования. После удовлетворения одной ведущей потребности доминирующее положение занимает другая наиважнейшая по социальной или биологической значимости потребность.
Принцип последовательного динамического взаимодействия предусматривает четкую последовательность смены деятельности нескольких взаимосвязанных ФС. Фактором, определяющим начало деятельности каждой последующей ФС, является результат деятельности предыдущей системы. Еще одним принципом организации взаимодействия ФС является принцип системного квантования жизнедеятельности. Например, в процессе дыхания можно выделить следующие системные «кванты» с их конечными результатами: вдох и поступление некоторого количества воздуха в альвеолы; диффузия О2 из альвеол в легочные капилляры и связывание О2 с гемоглобином; транспорт О2 к тканям; диффузия О2 из крови в ткани и СО2 в обратном направлении; транспорт СО2 к легким; диффузия СО2 из крови в альвеолярный воздух; выдох. Принцип системного квантования распространяется на поведение человека.
Таким образом, управление жизнедеятельностью организма путем организации ФС гомеостатического и поведенческого уровней обладает рядом свойств, позволяющих адекватно адаптировать организм к изменяющейся внешней среде. ФС позволяет реагировать на возмущающие воздействия внешней среды и на основе обратной аффектации перестраивать деятельность организма при отклонении параметров внутренней среды. Помимо этого, в центральных механизмах ФС формируется аппарат предвидения будущих результатов — акцептор результата действия, на основе которого происходит организация и инициация опережающих действительные события адаптивных актов, что существенно расширяет приспособительные возможности организма. Сравнение параметров достигнутого результата с афферентной моделью в акцепторе результатов действия служит основой для коррекции деятельности организма в плане получения именно тех результатов, которые наилучшим образом обеспечивают процесс адаптации.
Глава 4. НЕРВНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
МЕХАНИЗМЫ ДЕЯТЕЛЬНОСТИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Рефлекторный принцип регуляции функций
Основное положение рефлекторной теории заключается в утверждении, что деятельность организма есть закономерная рефлекторная реакция на стимул. Узловым моментом развития рефлекторной теории следует считать классический труд И. М. Сеченова (1863) «Рефлексы головного мозга», в котором впервые был провозглашен тезис о том, что все виды сознательной и бессознательной жизни человека представляют собой рефлекторные реакции. Рефлекс как универсальная форма взаимодействия организма и среды есть реакция организма, возникающая на раздражение рецепторов и осуществляемая с участием центральной нервной системы.
В естественных условиях рефлекторная реакция происходит при пороговом, надпороговом раздражении входа рефлекторной дуги — рецептивного поля данного рефлекса. Рецептивным полем называется определенный участок воспринимающей чувствительной поверхности организма с расположенными здесь рецепторными клетками, раздражение которых инициирует, запускает рефлекторную реакцию. Рецептивные поля разных рефлексов имеют определенную локализацию, рецепторные клетки — соответствующую специализацию для оптимального восприятия адекватных раздражителей (например, фоторецепторы располагаются в сетчатке; волосковые слуховые рецепторы — в спиральном (кортиевом) органе; проприорецепторы — в мышцах, в сухожилиях, в суставных полостях; вкусовые рецепторы на поверхности языка; обонятельные — в слизистой оболочке носовых ходов; болевые, температурные, тактильные рецепторы в коже и т. д.
Структурной основой рефлекса является рефлекторная дуга — последовательно соединенная цепочка нервных клеток, обеспечивающая осуществление реакции, или ответа, на раздражение. Рефлекторная дуга состоит из афферентного, центрального и эфферентного звеньев, связанных между собой синаптическими соединениями (рис. 4.1). Афферентная часть дуги начинается рецепторными образованиями, назначение которых заключается в трансформации энергии внешних раздражений в энергию нервного импульса, поступающего по афферентному звену дуги рефлекса в центральную нервную систему.
В зависимости от сложности структуры рефлекторной дуги различают моно- и полисинаптические рефлексы. В простейшем случае импульсы, поступающие в центральные нервные структуры по афферентным путям, переключаются непосредственно на эфферентную нервную клетку, т. е. в системе рефлекторной дуги имеется одно синаптическое соединение. Такая рефлекторная дуга называется моносинаптической (например, рефлекторная дуга сухожильного рефлекса в ответ на растяжение). Наличие в структуре рефлекторной дуги двух и более синаптических переключений (т. е. три и более нейронов), позволяет характеризовать ее как полисинаптическую.
Объяснение физиологических механизмов обучения, приобретенного навыка, коррекции выполненного ответа на основе принципа рефлекторной реакции неверно, неточно и потребовало существенного пересмотра классической схемы рефлекторной реакции, определяемой простой прямой связью: стимул à нервный центр à реакция.
Представление о рефлекторной реакции как о целесообразном ответе организма диктует необходимость дополнить рефлекторную дугу еще одним звеном — петлей обратной связи, призванной установить связь между реализованным результатом рефлекторной реакции и нервным центром, выдающим исполнительные команды. Обратная связь трансформирует открытую рефлекторную дугу в закрытую. Она может быть реализована разными способами: от исполнительной структуры к нервному центру (промежуточному или эфферентному двигательному нейрону), например, через возвратную аксонную коллатераль пирамидного нейрона коры больших полушарий или двигательной моторной клетки переднего рога спинного мозга. Обратная связь может обеспечиваться и нервными волокнами, поступающими к рецепторным структурам и управляющими чувствительностью рецепторных афферентных структур анализатора. Такая структура рефлекторной дуги превращает ее в самонастраивающийся нервный контур регуляции физиологической функции, совершенствуя рефлекторную реакцию и, в целом, оптимизируя поведение организма.
Классификации рефлексов. Существуют различные классификации рефлексов: по способам их вызывания, особенностям рецепторов, центральным нервным структурам их обеспечения, биологическому значению, сложности нейронной структуры рефлекторной дуги и т. д.
По способу вызывания различают безусловные рефлексы (категория рефлекторных реакций, передаваемых по наследству) и условные рефлексы (рефлекторные реакции, приобретаемые на протяжении индивидуальной жизни организма).
Различают экстероцептивные рефлексы — рефлекторные реакции, инициируемые раздражением многочисленных экстерорецепторов (болевые, температурные, тактильные и т. д.), интероцептивные рефлексы (рефлекторные реакции, запускаемые раздражением интероцепторов: хемо-, баро-, осморецепторов и т. д.), проприоцептивные рефлексы (рефлекторные реакции, осуществляемые в ответ на раздражение проприорецепторов мышц, сухожилий, суставных поверхностей и т. д.).
В зависимости от уровня активации части мозга дифференцируют спинномозговые, бульварные, мезенцефальные, диэнцефальные, кортикальные рефлекторные реакции.
По биологическому назначению рефлексы делят на пищевые, оборонительные, половые и т. д.
С учетом уровня эволюционного развития, совершенствования сложности нервного субстрата, обеспечивающего соответствующую рефлекторную реакцию, физиологического значения, уровня интегративной деятельности организма выделяют шесть основных видов рефлексов, или уровней рефлекторных реакций (А. Б. Коган):
Элементарные безусловные рефлексы, представлены простыми рефлекторными реакциями, осуществляемыми на уровне отдельных сегментов спинного мозга. Они имеют местное значение, вызываются локальным раздражением рецепторов данного сегмента тела и проявляются в виде локальных сегментарных сокращений поперечнополосатой мускулатуры. Элементарные безусловные рефлексы осуществляются по жестко детерминированным программам и имеют четкую определенную структурную основу в виде сегментарного аппарата спинного мозга, в результате такие рефлекторные реакции отличаются высокой степенью автоматизма и стереотипности. Функциональная роль этой категории рефлексов заключается в обеспечении простейших приспособительных реакций к внешним воздействиям местного значения, а также в приспособительных изменениях отдельных внутренних органов.
Координационные безусловные рефлексы представляют собой согласованные акты локомоторной деятельности или комплексные реакции вегетативных функциональных объединений внутренних органов. Эти рефлексы также вызываются раздражением определенных групп внешних или внутренних рецепторов, однако их эффект не ограничивается локальной реакцией путем последующей активации широкого класса экстеро-, интеро- и проприорецепторов, а формирует сложные координационные акты сокращения и расслабления, возбуждения или торможения деятельности ряда внутренних органов.
В физиологических механизмах реализации рефлекторных реакций этого типа значительное место занимает принцип обратной связи, обеспечиваемый соответствующими спинномозговыми структурами и осуществляющий антагонистическую, реципрокную иннервацию мышц-синергистов и антагонистов. Функциональное назначение координационных безусловных рефлексов — формирование на базе локальных элементарных безусловных рефлексов целостных, целенаправленных локомоторных актов или гомеостатических систем организма.
Интегративные безусловные рефлексы представляют собой дальнейший шаг в интеграции отдельных безусловных рефлексов, осуществляющих сложные двигательные локомоторные акты организма в тесной связи с вегетативным обеспечением, формируя тем самым комплексные поведенческие акты, имеющие определенное биологическое значение. Рефлекторные реакции этого типа инициируются такими биологически важными стимулами, как пищевые, болевые раздражители. Определяющим на входе этих рефлекторных актов становятся не физико-химические свойства стимулов, а в первую очередь их биологическое значение. Интегративные безусловные рефлексы всегда носят целостный системный характер, включая достаточно выраженные соматические и вегетативные компоненты. Их реализация оказывается весьма пластичной, тесно связанной со многими сильно развитыми проприоцептивными обратными связями, обеспечивающими точную коррекцию выполняемого сложного поведенческого акта в соответствии с изменениями в состоянии организма. Пример такой реакции — ориентировочная реакция. Биологическое значение последней заключается в перестройке организма, которая обеспечивает оптимальную подготовку к восприятию и быстрому анализу нового неизвестного сигнала в целях организации рационального ответа. Интегративные безусловные рефлексы требуют для своей реализации надсегментарных механизмов нервно-рефлекторной регуляции поведения организма. Эти рефлексы означают переход от сравнительно простых безусловных рефлексов к поведенческим актам.
Сложнейшие безусловные рефлексы (инстинкты) представляют собой видовые стереотипы поведения, организующиеся на базе интегративных рефлексов по генетически заданной программе. В качестве запускающих стереотипные поведенческие реакции раздражений выступают стимулы, имеющие отношение к питанию, защите, размножению и другим биологически важным потребностям организма.
Сложнейшие безусловные рефлексы образованы последовательными интегративными реакциями, построенными таким образом, что завершение одной реакции становится началом следующей. Адаптивность инстинктов усиливается благодаря наслоению на сложнейшие безусловные рефлексы условных, приобретаемых на ранних этапах онтогенеза. Нервный субстрат, ответственный за физиологические механизмы инстинктивного поведения, представляет иерархическую систему соподчиненных центров интегративных, координационных и элементарных безусловных рефлексов. Жесткая предопределенность инстинктивных реакций обусловлена этапной последовательностью актов инстинктивного поведения, ограничивающей сферу функционирования обратной связи от последующего этапа к предыдущему, уже реализованному. Инстинктивные реакции отражают исторический опыт вида. В субъективной сфере человека сложнейшие безусловные рефлексы проявляются в виде последовательных влечений и желаний, в сложной игре эмоций.
Элементарные условные рефлексы проявляются в интегративных реакциях, вызываемых ранее индифферентными раздражителями, приобретающими сигнальное значение в результате жизненного опыта или подкрепления их безусловными стимулами (сигналами), имеющими биологическое значение. Основным принципиальным отличием этой категории рефлекторных реакций является то, что они образуются в процессе индивидуальной жизни. Условнорефлекторные реакции образуются, усложняются, видоизменяются на протяжении всей жизни; наиболее простые из них формируются в раннем возрасте. Нервным субстратом, отвечающим за осуществление условнорефлекторных реакций, является филогенетически наиболее молодая структура головного мозга — кора больших полушарий. Многоканальность и взаимозаменяемость путей реализации условнорефлекторного механизма лежат в основе высокой пластичности и надежности условнорефлекторных реакций. В системе рефлекторных реакций появление условного рефлекса означает качественно новый скачок в приспособительном поведении высших животных и человека. Условнорефлекторные реакции дают возможность организму заблаговременно отвечать на приближающиеся жизненно важные ситуации. В психической сфере деятельности человека условные рефлексы закладывают начало ассоциативному способу мышления.
Сложные формы высшей нервной деятельности представлены психическими реакциями, возникающими на основе интеграции элементарных условных рефлексов и аналитико-синтетических механизмов абстрагирования. Абстрагирование от конкретного содержания безусловных подкрепляющих раздражителей обеспечивает возможность более полного и целостного восприятия окружающего мира, адекватного прогнозирования и программирования поведения. В качестве вызывающих подобные реакции стимулов обычно выступают сложные комплексные раздражители. Часто такие рефлекторные реакции имеют усеченную рефлекторную дугу (отсутствует эфферентное звено рефлекторной дуги). Сложные формы высшей нервной деятельности оказываются связанными с синтетическими процессами, обеспечивающими целостные субъективные образы внешнего мира, целенаправленные программы поведения, различные проявления абстрактной мыслительной деятельности человека (психонервная деятельность, рассудочное мышление, функции второй сигнальной системы).
Глава 4. НЕРВНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
МЕХАНИЗМЫ ДЕЯТЕЛЬНОСТИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Торможение в центральной нервной системе
Интегративная и координационная деятельность центральных нервных образований осуществляется при обязательном участии тормозных процессов.
Торможение в центральной нервной системе — активный процесс, проявляющийся внешне в подавлении или в ослаблении процесса возбуждения и характеризующийся определенной интенсивностью и длительностью.
Торможение в норме неразрывно связано с возбуждением, является его производным, сопутствует возбудительному процессу, ограничивая и препятствуя чрезмерному распространению последнего. При этом торможение часто ограничивает возбуждение и вместе с ним формирует сложную мозаику активированных и заторможенных зон в центральных нервных структурах. Формирующий эффект тормозного процесса развивается в пространстве и во времени. Торможение — врожденный процесс, постоянно совершенствующийся в течение индивидуальной жизни организма.
При значительной силе фактора, вызвавшего торможение, оно может распространяться на значительное пространство, вовлекая в тормозной процесс большие популяции нервных клеток.
История развития учения о тормозных процессах в центральной нервной системе начинается с открытия И. М. Сеченовым эффекта центрального торможения (химическое раздражение зрительных бугров тормозит простые спинномозговые безусловные реакции). Вначале предположение о существовании специфических тормозных нейронов, обладающих способностью оказывать тормозные влияния на другие нейроны, с которыми имеются синаптические контакты, диктовалось логической необходимостью для объяснения сложных форм координационной деятельности центральных нервных образований. Впоследствии это предположение нашло прямое экспериментальное подтверждение (Экклс, Реншоу), когда было показано существование специальных вставочных нейронов, имеющих синаптические контакты с двигательными нейронами. Активация этих вставочных нейронов закономерно приводила к торможению двигательных нейронов. В зависимости от нейронного механизма, способа вызывания тормозного процесса в ЦНС различают несколько видов торможения: постсинаптическое, пресинаптическое, пессимальное.
Постсиналтическое торможение — основной вид торможения, развивающийся в постсинаптической мембране аксосоматических и аксодендритических синапсов под влиянием активации тормозных нейронов, в концевых разветвлениях аксонных отростков которых освобождается и поступает в синаптическую щель тормозной медиатор. Тормозной эффект таких нейронов обусловливается специфической природой медиатора — химического переносчика сигнала с одной клетки на другую. Наиболее распространенным тормозным медиатором является гамма-аминомасляная кислота (ГАМК). Химическое действие ГАМК вызывает в постсинаптической мембране эффект гиперполяризации в виде тормозных постсинаптических потенциалов (ТПСП), пространственно-временная суммация которых повышает уровень мембранного потенциала (гиперполяризация), приводит к урежению или полному прекращению генерации распространяющихся ПД.
Возвратным торможением называется угнетение (подавление) активности нейрона, вызываемое возвратной коллатералью аксона нервной клетки. Так, мотонейрон переднего рога спинного мозга прежде чем покинуть спинной мозг дает боковую (возвратную) ветвь, которая возвращается назад и заканчивается на тормозных нейронах (клетки Реншоу). Аксон последней заканчивается на мотонейронах, оказывая на них тормозное действие.
Пресинаптическое торможение развертывается в аксоаксональных синапсах, блокируя распространение возбуждения по аксону. Пресинаптическое торможение часто выявляется в структурах мозгового ствола, в спинном мозге
Пессимальное торможение представляет собой вид торможения центральных нейронов. Оно наступает при высокой частоте раздражения. В первый момент возникает высокая частота ответного возбуждения. Через некоторое время стимулируемый центральный нейрон, работая в таком режиме, переходит в состояние торможения.
Глава 4. НЕРВНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
МЕХАНИЗМЫ ДЕЯТЕЛЬНОСТИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Свойства нервных центров
Рефлекторная деятельность организма во многом определяется общими свойствами нервных центров.
Нервный центр — совокупность структур центральной нервной системы, координированная деятельность которых обеспечивает регуляцию отдельных функций организма или определенный рефлекторный акт. Представление о структурно-функциональной основе нервного центра обусловлено историей развития учения о локализации функций в центральной нервной системе. На смену старым теориям об узкой локализации, или эквипотенциальности, высших отделов головного мозга, в частности коры большого мозга, пришло современное представление о динамической локализации функций, основанное на признании существования четко локализованных ядерных структур нервных центров и менее определенных рассеянных элементов анализаторных систем мозга. При этом с цефализацией нервной системы растут удельный вес и значимость рассеянных элементов нервного центра, внося существенные различия в анатомических и физиологических границах нервного центра. В результате функциональный нервный центр может быть локализован в разных анатомических структурах. Например, дыхательный центр представлен нервными клетками, расположенными в спинном, продолговатом, промежуточном мозге, в коре большого мозга.
Нервные центры имеют ряд общих свойств, что во многом определяется структурой и функцией синаптических образований.
Односторонность проведения возбуждения. В рефлекторной дуге, включающей нервные центры,
процесс возбуждения распространяется в одном направлении (от входа, афферентных путей к выходу, эфферентным путям).
Иррадиация возбуждения. Особенности структурной организации центральных нейронов, огромное
число межнейронных соединений в нервных центрах существенно модифицируют (изменяют) направление распространения процесса возбуждения в зависимости от силы раздражителя и функционального состояния центральных нейронов. Значительное увеличение силы раздражителя приводит к расширению области вовлекаемых в процесс возбуждения центральных нейронов — иррадиации возбуждения.
Суммация возбуждения. В работе нервных центров значительное место занимают процессы пространственной и временной суммации возбуждения, основным нервным субстратом которой является постсинаптическая мембрана. Процесс пространственной суммации афферентных потоков возбуждения облегчается наличием на мембране нервной клетки сотен и тысяч синаптических контактов. Процессы временной суммации обусловлены суммацией ВПСП на постсинаптической мембране.
Наличие синаптической задержки. Время рефлекторной реакции зависит в основном от двух факторов: скорости движения возбуждения по нервным проводникам и времени распространения возбуждения с одной клетки на другую через синапс. При относительно высокой скорости распространения импульса по нервному проводнику основное время рефлекса приходится на синаптическую передачу возбуждения (синаптическая задержка). В нервных клетках высших животных и человека одна синаптическая задержка примерно равна 1 мс. Если учесть, что в реальных рефлекторных дугах
имеются десятки последовательных синаптических контактов, становится понятной длительность большинства рефлекторных реакций — десятки миллисекунд.
Высокая утомляемость. Длительное повторное раздражение рецептивного поля рефлекса приводит к ослаблению рефлекторной реакции вплоть до полного исчезновения, что называется утомлением. Этот процесс связан с деятельностью синапсов — в последних наступает истощение запасов медиатора, уменьшаются энергетические ресурсы, происходит адаптация постсинаптического рецептора к медиатору.
Тонус. Тонус, или наличие определенной фоновой активности нервного центра, определяется тем, что в покое в отсутствие специальных внешних раздражений определенное количество нервных клеток находится в состоянии постоянного возбуждения, генерирует фоновые импульсные потоки. Даже во сне в высших отделах мозга остается некоторое количество фоновоактивных нервных клеток, формирующих «сторожевые пункты» и определяющих некоторый тонус соответствующего нервного центра.
Пластичность. Функциональная возможность нервного центра существенно модифицировать картину осуществляемых рефлекторных реакций. Поэтому пластичность нервных центров тесно связана с изменением эффективности или направленности связей между нейронами.
Конвергенция. Нервные центры высших отделов мозга являются мощными коллекторами, собирающими разнородную афферентную информацию. Количественное соотношение периферических рецепторных и промежуточных центральных нейронов (10:1) предполагает значительную конвергенцию («сходимость») разномодальных сенсорных посылок на одни и те же центральные нейроны. На это указывают прямые исследования центральных нейронов: в нервном центре имеется значительное количество поливалентных, полисенсорных нервных клеток, реагирующих на разномодальные стимулы (свет, звук, механические раздражения и т. д.). Конвергенция на клетках нервного центра разных афферентных входов предопределяет важные интегративные, перерабатывающие информацию функции центральных нейронов, т. е. высокий уровень интеграционных функций. Конвергенция нервных сигналов на уровне эфферентного звена рефлекторной дуги определяет физиологический механизм принципа «общего конечного пути» по Ч. Шеррингтону.
Интеграция в нервных центрах. Важные интегративные функции клеток нервных центров ассоциируются с интегративными процессами на системном уровне в плане образования функциональных объединений отдельных нервных центров в целях осуществления сложных координированных приспособительных целостных реакций организма (сложные адаптивные поведенческие акты).
Свойство доминанты. Доминантным называется временно господствующий в нервных центрах очаг (или доминантный центр) повышенной возбудимости в центральной нервной системе. По А.А.Ухтомскому, доминантный нервный очаг характеризуется такими свойствами, как повышенная возбудимость, стойкость и инертность возбуждения, способность к суммированию возбуждения.
В доминантном очаге устанавливается определенный уровень стационарного возбуждения, способствующий суммированию ранее подпороговых возбуждений и переводу на оптимальный для данныхусловий ритм работы, когда этот очаг становится наиболее чувствительным. Доминирующее значение такого очага (нервного центра) определяет его угнетающее влияние на другие соседние очаги возбуждения. Доминантный очаг возбуждения «притягивает» к себе возбуждение других возбужденных зон (нервных центров). Принцип доминанты определяет формирование главенствующего (активирующего) возбужденного нервного центра в тесном соответствии с ведущими мотивами, потребностями организма в конкретный момент времени.
11. Цефализация нервной системы. Основная тенденция в эволюционном развитии нервной системы проявляется в перемещении, сосредоточении функции регуляции и координации деятельности организма в головных отделах ЦНС. Этот процесс называется цефализацией управляющей функции нервной системы. При всей сложности складывающихся отношений между старыми, древними и эволюционно новыми нервными образованиями стволовой части мозга общая схема взаимных влияний может быть представлена следующим образом: восходящие влияния (от нижележащих «старых» нервных структур к вышележащим «новым» образованиям) преимущественно носят возбуждающий стимулирующий характер, нисходящие (от вышележащих «новых» нервных образований к нижележащим «старым» нервным структурам) носят угнетающий тормозной характер. Эта схема согласуется с представлением о росте в процессе эволюции роли и значения тормозных процессов в осуществлении сложных интегративных рефлекторных реакций.
Глава 4. НЕРВНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
МЕХАНИЗМЫ ДЕЯТЕЛЬНОСТИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Принципы интеграции и координации в деятельности центральной нервной системы
В реализации информационно-управляющей функции нервной системы значительная роль принадлежит процессам интеграции и координации деятельности отдельных нервных клеток и нейронных ансамблей, которые основаны на особенностях взаимодействия информационных потоков на уровне нервных клеток и рефлекторных дуг. Конструктивные особенности афферентных, промежуточных (центральных) и эфферентных нейронов обеспечивают широкий диапазон иррадиации и концентрации возбуждения на основе двух кардинальных принципов: дивергенции и конвергенции. Дивергенцией называется способность нервной клетки устанавливать многочисленные синаптические связи с различными нервными клетками. Благодаря этому одна нервная клетка может участвовать в нескольких различных реакциях, передавать возбуждение значительному числу других нейронов, которые могут возбудить большее количество нейронов, обеспечивая широкую иррадиацию возбудительного процесса в центральных нервных образованиях.
Процессы конвергенции заключаются в схождении различных импульсных потоков от нескольких нервных клеток к одному и тому же нейрону (см. раздел 4.1.4). Процесс конвергенции характерен не только для однотипных нервных клеток. Например, на мотонейронах спинного мозга, кроме первичных афферентных волокон, конвергируют волокна различных нисходящих трактов от супраспинальных и собственно спинальных центров, а также от возбуждающих и тормозных вставочных промежуточных нейронов. В результате мотонейроны спинного мозга выполняют функцию общего конечного пути для многочисленных нервных образований, включая и надсегментный аппарат головного мозга, имеющих отношение к регуляции двигательной функции.
В координационной деятельности центральных нервных образований значительная роль взаимодействия рефлексов, которая проявляется в различных эффектах (в облегчении, или суммации, и в угнетении, или подавлении, возбуждения).
Примером координационного взаимодействия рефлексов является реципрокная иннервация мышц-антагонистов. Известно, что сгибание или разгибание конечностей осуществляется благодаря согласованной работе двух функционально антагонистических мышц: сгибателей и разгибателей. Координация обеспечивается организацией антагонистических отношений между мотонейронами сгибателей и разгибателей, иннервирующих соответствующие мышцы. Реципрокные функциональные отношения складываются в сегментарных структурах спинного мозга благодаря включению в дугу спинномозгового рефлекса дополнительного элемента — специального тормозного нейрона (клетка Реншоу). Схематически в общем виде этот механизм выглядит следующим образом: сигнал от афферентного звена через обычную вставочную (промежуточную) нервную клетку вызывает возбуждение мотонейрона, иннервирующего мышцу-сгибатель, а через дополнительную клетку Реншоу тормозит мотонейрон, иннервирующий мышцу-разгибатель. Так происходит координированное сгибание конечности; напротив, при выполнении разгибания конечности возбуждается мотонейрон мышцы-разгибателя, а через вставочную клетку Реншоу тормозится, угнетается мотонейрон мышцы-сгибателя (рис. 4.3). Таким образом акт сгибания конечности, представляющий собой работу мышц-антагонистов, обеспечивается формированием реципрокных антагонистических отношений между нервными центрами соответствующих мышц благодаря наличию специальных тормозных нейронов — клеток Реншоу.
В центральной нервной системе весьма распространены интегративные феномены — посттетаническая потенциация, окклюзия, облегчение.
Феномен посттетанической потенциации проявляется следующим образом. Раздражая стимулами редкой частоты афферентный нерв, можно получить некоторый рефлекс определенной интенсивности. Если затем этот нерв в течение некоторого времени подвергать высокочастотному ритмическому раздражению (300—400 стимулов в секунду), то повторное редкое ритмическое раздражение приведет к резкому усилению реакции
Если два нервных центра рефлекторных реакций имеют частично перекрываемые рецептивные поля, то при совместном раздражении обоих рецептивных полей реакция будет меньше, чем арифметическая сумма реакций при изолированном раздражении каждого из рецептивных полей — феномен окклюзии (рис. 4.5). В ряде случаев вместо такого ослабления реакции при совместном раздражении рецептивных полей двух рефлексов можно наблюдать феномен облегчения (т. е. суммарная реакция выше суммы реакции при изолированном раздражении этих рецептивных полей). Это результат того, что часть общих для обоих рефлексов нейронов при изолированном раздражении оказывает подпороговый эффект для вызывания рефлекторных реакций. При совместном раздражении они суммируются и достигают пороговой силы, в результате конечная реакция оказывается больше суммы изолированных реакций.
Глава 4. НЕРВНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
МЕХАНИЗМЫ ДЕЯТЕЛЬНОСТИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Нейронные комплексы и их роль в деятельности центральной нервной системы
Концепция организации и самоорганизации в строении и функций нервной системы получила наибольшее развитие в представлениях о модульной (ансамблевой) конструкции нервной системы как принципиальной основы построения функциональных систем мозга. Хотя простейшей структурной и функциональной единицей нервной системы является нервная клетка, многочисленные данные современной нейрофизиологии подтверждают тот факт, что сложные функциональные «узоры» в центральных нервных образованиях определяются эффектами скоординированной активности в отдельных популяциях (ансамблях) нервных клеток.
Образования головного мозга состоят из повторяющихся локальных нейронных сетей модулей, которые варьируют от структуры к структуре по числу клеток, внутренним связям и способу обработки информации. Каждый модуль, или нейронный ансамбль, представляет собой совокупность локальных нейронных сетей, которая обрабатывает информацию, передает ее со своего входа на выход, подвергает трансформации, определяемой общими свойствами структуры и ее внешними связями. Один модуль может входить в состав различных функциональных образований.
Группирование нейронов в ансамбли нервных клеток для совместного выполнения функций следует рассматривать как проявление кооперативного способа деятельности. Основным функциональным признаком ансамблевой организации является локальный синергизм реакций нейронов центральной ядерной структуры ансамбля, окруженной зоной заторможенных и нереагирующих нейронов (А. Б. Коган, О. Г. Чораян). Размеры группировок нейронов в горизонтальной плоскости в среднем достигают диаметра 100—150 мкм, что соответствует размерам клеточных объединений, выявляемых по функциональным показателям синергичности возбудительных реакций на адекватное раздражение их рецептивных полей. Размеры зоны синаптических окончаний вторичного специфического афферентного волокна в корковых структурах (100—150 мкм) близок к пространственным характеристикам элементарного нейронного ансамбля. Примерно такие же размеры имеет и сфера терминальных разветвлений отдельного неспецифического волокна, но общая зона всех ветвей неспецифического волокна образует сферу диаметром 600— 700 мкм, что соответствует размерам зоны ветвления первичного специфического афферентного волокна.
Схема активации нейронного ансамбля может быть представлена следующим образом. Сигналы, поступающие по первичным специфическим и неспецифическим афферентам, активируют вначале обширную зону, вовлекая в процесс возбуждения группу нейронных ансамблей. Более дробная конфигурация нейронных группировок в зоне диаметром 100—150 мкм формируется под влиянием вторичных афферентных волокон, несущих сигналы внутрицентрального взаимодействия (рис. 4.6). Из множества элементарных нейронных ансамблей образуется центральная мозаика активности, определяющая постоянно меняющийся «узор» возбуждения и торможения в нервном центре. Таким образом, ансамблевая конструкция центральных проекционных зон анализатора в коре большого мозга представляется как результат двух физиологических механизмов: 1) мощной активации большой зоны центральных нейронов, связанных с терминалами афферентных волокон; 2) центральными нейронами, в функциональном отношении аналогичными клеткам Реншоу в спинном мозге, препятствующими широкому растеканию центрального возбуждения путем формирования тормозной каемки вокруг возбужденных нервных клеток. Разнообразие «узоров» возбуждения и торможения в центральной мозаике нейрональной активности формируется из элементарных микроочагов возбуждения, которые образуют фундамент иерархической конструкции клеточных систем мозга.
Принципиальным моментом ансамблевой концепции работы мозга является утверждение, что на каждом этапе переработки информации в качестве функциональной единицы выступает не отдельно взятая нервная клетка, а внутренне интегрированное клеточное объединение — нейронный ансамбль, основными характеристиками которого является: а) локальный синергизм реакции нейронов центральной зоны; б) наличие тормозной окантовки, образованной клетками с тормозными реакциями на данное раздражение, окружающими центральную зону клеток с возбудительными реакциями; в) наличие определенного числа нейронов со стабильными ответами (обычно они расположены в центральной ядерной зоне ансамбля) при значительно большем числе клеток с вариабельными параметрами импульсного ответа на адекватное афферентное раздражение. Элементарные нейронные ансамбли как функциональные единицы рабочих механизмов мозга играют роль своего рода «кирпичиков», из которых формируются более сложные блоки и конструкции мозга.
Наблюдающаяся структурная и функциональная избыточность ансамблевой модульной конструкции центральных нервных образований как следствие ансамблевой организации ассоциируется со значительной информационной избыточностью сенсорных посылок, распространяющихся по нейронным системам мозга. Избыточность нейронных элементов и межнейронных связей в ансамбле — характерная черта структурно-функциональной организации центральных нервных образований — приводит к мультифункциональности, которой обусловлены пластичность и высокие компенсаторные способности нервных механизмов.
Когда индивидуальное участие нейрона в данной реакции становится не обязательным, а вероятностным и возможна относительная взаимозаменяемость элементов, повышаются надежность нервного механизма управления и связи в организме. Подвижная динамическая структура нейронных ансамблей, формируемая вероятностным участием в них отдельных нервных клеток, обусловливает большую гибкость и легкость перестроек межнейронных связей; эти перестройки объясняют высокую пластичность, характерную для нервных механизмов высших отделов мозга.
Вероятностные нейронные ансамбли образуются структурно-функциональными блоками нервных клеток, воспринимающих и перерабатывающих идентичную афферентную информацию. Эти ансамбли составляют основу функциональной мозаики процессов центрального возбуждения и торможения. Мозаика вероятностных нейронных ансамблей на всех уровнях конструкции нервного контура управления, обеспечивающего разные формы целенаправленного поведения, образует функциональную систему мозга.
Глава 4. НЕРВНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
МЕХАНИЗМЫ ДЕЯТЕЛЬНОСТИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Гематоэнцефалический барьер и его функции
Организм человека и высших животных обладает рядом специфических физиологических систем, обеспечивающих приспособление (адаптацию) к постоянно изменяющимся условиям существования. Этот процесс тесно связан с необходимостью обязательного сохранения постоянства существенных физиологических параметров, внутренней среды организма, физико-химического состава тканевой жидкости межклеточного пространства.
Среди гомеостатических приспособительных механизмов, призванных защитить органы и ткани от чужеродных веществ и регулировать постоянство состава тканевой межклеточной жидкости, ведущее место занимает гематоэнцефалический барьер. По определению Л. С. Штерн, гематоэнцефалический барьер объединяет совокупность физиологических механизмов и соответствующих анатомических образований в центральной нервной системе, участвующих в регулировании состава цереброспинальной жидкости (ЦСЖ).
В представлениях о гематоэнцефалическом барьере в качестве основных положений подчеркивается следующее: 1) проникновение веществ в мозг осуществляется главным образом не через ликворные пути, а через кровеносную систему на уровне капилляр — нервная клетка; 2) гематоэнцефалический барьер является в большей степени не анатомическим образованием, а функциональным понятием, характеризующим определенный физиологический механизм. Как любой существующий в организме физиологический механизм, гематоэнцефалический барьер находится под регулирующим влиянием нервной и гуморальной систем; 3) среди управляющих гематоэнцефалическим барьером факторов ведущим является уровень деятельности и метаболизма нервной ткани.
Гематоэнцефалический барьер регулирует проникновение из крови в мозг биологически активных веществ, метаболитов, химических веществ, воздействующих на чувствительные структуры мозга, препятствует поступлению в мозг чужеродных веществ, микроорганизмов, токсинов.
Основной функцией, характеризующей гематоэнцефалический барьер, является проницаемость клеточной стенки. Необходимый уровень физиологической проницаемости, адекватный функциональному состоянию организма, обусловливает динамику поступления в нервные клетки мозга физиологически активных веществ.
Функциональная схема гематоэнцефалического барьера включает в себя наряду с гистогематическим барьером нейроглию и систему ликворных пространств (Я. А. Росин) (схема 4.1). Гистогематический барьер имеет двойную функцию: регуляторную и защитную. Регуляторная функция обеспечивает относительное постоянство физических и физико-химических свойств, химического состава, физиологической активности межклеточной среды органа в зависимости от его функционального состояния. Защитная функция гистогематического барьера заключается в защите органов от поступления чужеродных или токсичных веществ эндо- и экзогенной природы.
Ведущим компонентом морфологического субстрата гематоэнцефалического барьера, обеспечивающим его функции, является стенка капилляра мозга. Существуют два механизма проникновения вещества в клетки мозга: через цереброспинальную жидкость, которая служит промежуточным звеном между кровью и нервной или глиальной клеткой, которая выполняет питательную функцию (так называемый ликворный путь), и через стенку капилляра. У взрослого организма основным путем движения вещества в нервные клетки является гематогенный (через стенки капилляров); ликворный путь становится вспомогательным, дополнительным.
Проницаемость гематоэнцефалического барьера зависит от функционального состояния организма, содержания в крови медиаторов, гормонов, ионов. Повышение их концентрации в крови приводит к снижению проницаемости гематоэнцефалического барьера для этих веществ.
Функциональная система гематоэнцефалического барьера представляется важным компонентом нейрогуморальной регуляции. В частности, через гематоэнцефалический барьер реализуется принцип обратной химической связи в организме. Именно таким образом осуществляется механизм гомеостатической регуляции состава внутренней среды организма.
Регуляция функций гематоэнцефалического барьера осуществляется высшими отделами ЦНС и гуморальными факторами. Значительная роль в регуляции отводится гипоталамо-гипофизарной адреналовой системе. В нейрогуморальной регуляции гематоэнцефалического барьера важное значение имеют обменные процессы, в частности в ткани мозга. При различных видах церебральной патологии, например травмах, различных воспалительных поражениях ткани мозга, возникает необходимость искусственного снижения уровня проницаемости гематоэнцефалического барьера. Фармакологическими воздействиями можно увеличить или уменьшить проникновение в мозг различных веществ, вводимых извне или циркулирующих в крови.
Глава 4. НЕРВНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
МЕХАНИЗМЫ ДЕЯТЕЛЬНОСТИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Цереброспинальная жидкость
Цереброспинальная жидкость (син.: ликвор, спинномозговая жидкость) — прозрачная бесцветная жидкость, заполняющая полости желудочков мозга, субарахноидальное пространство головного мозга и спинномозговой канал, периваскулярные и перицеллюлярные пространства в ткани мозга. Цереброспинальная жидкость выполняет питательные функции, а также определяет величину внутримозгового давления. Состав цереброспинальной жидкости формируется в процессе обмена веществ между мозгом, кровью и тканевой жидкостью, включая все компоненты ткани мозга. В цереброспинальной жидкости содержится ряд биологически активных соединений: гормоны гипофиза и гипоталамуса, ГАМК, АХ, норадреналин, дофамин, серотонин, малатонин, продукты их метаболизма.
Среди клеток цереброспинальной жидкости преобладают лимфоциты (более 60% от общего числа клеток) — в норме в 1 мкл спинномозговой жидкости содержится 1,5 клетки. Химический состав цереброспинальной жидкости очень близок к таковому крови: 89— 90% воды, 10—11% сухого остатка, содержащего органические и неорганические вещества, участвующие в метаболизме мозговой ткани. Общий белок цереброспинальной жидкости содержит до 30 различных фракций; основную часть его формируют миелин и образующиеся при его распаде промежуточные продукты, гликопептиды, липопротеины, полиамины, белок S-100. Цереброспинальная жидкость содержит лизоцим, ферменты (кислая и щелочная фосфатазы, рибонуклеазы, лактатдегидрогеназа, ацетилхолинэстераза, пептидазы и др.).
В клинической практике важную диагностическую значимость имеет белковый коэффициент Кафки цереброспинальной жидкости — отношение количества глобулинов к альбуминам. В норме он составляет 0,2—0,3.
К основным ликвороносным путям относятся боковые желудочки, III и IV желудочки головного мозга, водопровод среднего мозга, сильвиев водопровод, цистерны головного и спинного мозга. Система ликворообращения мозга включает три основных звена: ликворопродукцию, ликвороциркуляцию и отток ликвора.
Продукция цереброспинальной жидкости осуществляется в основном сосудистыми сплетениями желудочков мозга путем фильтрации из плазмы крови. В образовании цереброспинальной жидкости принимают участие структурные элементы мозга благодаря возможности диффузии межклеточной жидкости через эпендиму в желудочки головного мозга и через межклеточные пространства к поверхности мозга. В продукции цереброспинальной жидкости принимают участие и клетки мозговой ткани (нейроны и глия). В нормальных условиях экстраваскулярная продукция цереброспинальной жидкости весьма незначительна.
Путь постоянной циркуляции цереброспинальной жидкости схематически выглядит следующим образом: из боковых желудочков мозга через межжелудочковое отверстие (отверстие Монро) она поступает в III желудочек, затем через водопровод среднего мозга — в IV желудочек, откуда большая часть жидкости через срединную апертуру (отверстие Мажанди) и латеральные апертуры (отверстия Лушки) переходит в цистерны основания мозга, достигает борозды среднего мозга (сильвиева борозда) и поднимается в субарахноидальное пространство полушарий большого мозга. Циркуляция цереброспинальной жидкости определяется градиентом гидростатического давления в ликворных путях, пространствах мозга, обусловленного пульсацией внутричерепных кровеносных сосудов, изменениями венозного давления и положения тела в пространстве.
Отток цереброспинальной жидкости преимущественно (на 30— 40%) происходит через арахноидальное пространство в продольный синус (часть венозной системы головного мозга). Движущим фактором такого перемещения цереброспинальной жидкости является градиент гидростатического давления ее и венозной крови. Давление цереброспинальной жидкости в норме превышает венозное в верхнем продольном синусе на 15—20 мм вод. ст. Около 10% жидкости оттекает через сосудистое сплетение желудочков мозга, от 5 до 30% — в лимфатическую систему через периневральные пространства черепных и спинномозговых нервов. Некоторое количество жидкости резорбируется эпендимой желудочков мозга и сосудистыми сплетениями.
Общий объем циркуляции цереброспинальной жидкости у взрослого человека в норме составляет 90—200 мл, в среднем 140 мл. В сутки вырабатывается около 500 мл цереброспинальной жидкости, обновление ее происходит примерно 4—8 раз в сутки. Значительные колебания в скорости обновления цереброспинальной жидкости зависят от суточного режима питания, водного режима, колебаний активности физиологических процессов в организме, физиологической нагрузки на ЦНС и др.
Скорость образования цереброспинальной жидкости может значительно возрастать при развитии патологических процессов (воспалительные процессы, черепномозговые травмы, субарахноидальные кровоизлияния, операции по поводу опухоли мозга и т. д.).
Элементы кибернетики нервной системы
Нейрокибернетика (кибернетика нервной системы) — наука, изучающая процессы управления и связи в нервной системе. Такое определение предмета и задач кибернетики нервной системы позволяет выделить три составных компонента (раздела) ее: организация, управление и информационная деятельность.
В сложных полифункциональных интегративных системах мозга невозможно раздельное функционирование элементов организации, управления и информационной деятельности, они тесно связаны и взаимообусловлены. Организация нервной системы во многом предопределяет механизмы управления и эффективности передачи и переработки информации. Управление модифицирует механизмы организации и самоорганизации, обеспечивает эффективность и надежность информационной функции системы. Информационная деятельность является обязательным условием совершенствования процесса организации, управления как оперативный прием эффективного воздействия и целенаправленного видоизменения.
Организация. В центре внимания теории организации и самоорганизации в нейрокибернетике лежит представление о системных свойствах конструкций мозга на разных морфологических и эволюционных уровнях конструкции нервной системы. Ведущим свойством системы является организация. Система — совокупность элементов, где конечный результат кооперации проявляется не в виде суммы эффектов составляющих элементы, а в виде произведения эффектов, т. е. системность как характерное свойство организованной сложности предполагает неаддитивное сложение функций отдельных компонентов. Объединение двух и более элементов в системе рождает новое качество, которое не может быть выражено через качество составляющих компонентов.
Отдельный нейрон является носителем свойств, позволяющих ему интегрировать влияние других нейронов, строить свою активность на основании оценки результатов интеграции. С другой стороны, на основе таких свойств происходит объединение индивидуальных нейронов в системы, обладающие новыми свойствами, отсутствующими у входящих в их состав единиц. Характерной чертой таких систем является то, что активность каждого составного элемента в них определяется не только влияниями, поступающими по прямым афферентным путям каждого элемента, но и состоянием других элементов системы. Свойство системности в нервных образованиях возникает тогда, когда деятельность каждой нервной клетки оказывается функцией не только непосредственно поступившего к ней сигнала, но и функцией тех процессов, которые происходят в остальных клетках нервного центра (П. Г. Костюк).
Оптимальная организация нервных конструкций обычно сочетается со значительной структурой или функциональной избыточностью, которой принадлежит решающая роль в обеспечении пластичности и надежности биологической системы.
Нервная система животных и человека — самая совершенная по структуре система, разнообразие форм и размеров клеток которой не имеет аналога ни в какой другой физиологической системе биологического организма. Все многообразие и сложность форм нервных клеток в разных структурах и органах есть результат и основа богатого разнообразия функций элементов ведущей регуляторной системы организма. Часто наблюдаемые петлеобразные структуры в архитектонике волокнистых структур мозга (боковые и возвратные ветви аксонных отростков), обеспечивающих возможность циркуляторного прохождения информации, очевидно, выполняют функции механизма обратной связи, играющей столь важную роль в кибернетике нервной системы.
Важным моментом организации и самоорганизации служит системообразующий фактор — результат действия (П.К.Анохин). Реальной физиологической системой нейронов является комплекс нервных клеток, у которых взаимодействие и взаимоотношения приобретают характер взаимодействия элементов на получение фиксированного полезного результата (см. раздел 3.3).
Управление. Суть процесса управления заключается в том, что из множества возможных воздействий отбираются и реализуются те, которые направлены на поддержание, обеспечение рассматриваемой функции органа. Управление представляет собой информационный процесс, предусматривающий обязательность контроля за поведением объекта благодаря кольцевой, или круговой, передаче сигналов. Это предусматривает два вида передачи информации: по цепи управления от регулятора к объекту и в обратном направлении — от объекта к регулятору, при помощи обратной связи, по которой поступает информация о фактическом состоянии управляемого объекта.
Обратная связь бывает двух видов: положительной и отрицательной. В случае положительной обратной связи сигналы, поступающие на вход системы по цепи обратной связи, действуют в том же направлении, что и основные сигналы (воздействие среды). Положительная обратная связь ведет не к устранению, а к усилению рассогласования в системе. Отрицательная обратная связь обеспечивает выдачу управляемому объекту со стороны управляющего устройства команд, направленных на ликвидацию рассогласования действия системы (отклонений параметров системы от заданной программы). Стабилизирующая роль отрицательной обратной связи проявляется в том, что дополнительные сигналы, поступающие на вход системы по цепи обратной связи, действуют на систему в направлении, противоположном основному воздействию на объект.
В нейронных системах мозга встречаются два типа регулирования: управление по отклонению и управление по возмущению.
При управлении по отклонению, или по рассогласованию (величина ошибки), в качестве запускающего воздействия служит само отклонение регулируемой величины. В этом случае независимо от причины рассогласования возникшее отклонение вызывает регуляторные воздействия, направленные на его ликвидацию. Если этого окажется недостаточным для устранения эффекта возмущающего стимула, система мобилизует дополнительные механизмы обеспечения гомеостаза. Такой способ регулирования является наиболее простым и встречается в основном в примитивных формах организации нервной системы, на низших уровнях ее конструкции.
При управлении по возмущению регулирование осуществляется в ответ на внешний возмущающий сигнал до возникновения существенных отклонений в системе. Это более прогрессивный экономичный способ регуляции, свойственный высоким формам организации нервной системы.
Информационная функция. Ведущая роль нервной системы в организме определяется ее управляющей функцией по отношению к другим органам и тканям, обеспечиваемой благодаря способности воспринимать и перерабатывать информацию в целях оптимального приспособления организма к стохастической внешней среде. В процессе эволюционного филогенетического совершенствования нервных структур как ведущей информационной системы организма конструктивные особенности мозга определяют высокую адекватность (оптимальность) его коммуникационных систем: на мульти-клеточном уровне центральные нервные образования вместе с рецепторами и эффекторами составляют информационное поле с богатейшими возможностями для обработки сигналов.
Основным носителем информации в нервных клетках являются импульсные потоки, состоящие из отдельных импульсных сигналов стандартной амплитуды — распространяющихся потенциалов действия.
Центральным моментом в информационной деятельности нервных структур является кодирование, суть которого составляет процесс преобразования сообщения из одной формы в другую. Трансформированная в рецепторах информация подвергается в организме многократным дальнейшим превращениям на разных стадиях и уровнях организации нервной системы. Тонкая электрохимическая физиология рецепторов и синаптических соединений характеризует физический субстрат элементарных информационных превращений. В качестве кодирующих информацию элементов в самом импульсном потоке может быть любое статистическое измерение, характеризуемое определенным законом изменения в связи с различной интенсивностью раздражения.
В деятельности нервной системы значительное место занимают способы, методы пространственного кодирования информации, обеспечивающие высокую экономичность передачи информации о пространственном расположении, характеристике стимулов. Формы пространственного кодирования информации в дополнение к различным видам временного кодирования (интервальное, частотное и др.) существенно повышают информационную емкость нервных структур.
Сравнение суммарного информационного потока, поступающего в живой организм через органы чувств (3*109 бит/с) с количеством информации, необходимой для принятия целесообразного решения (20—25 бит/с), указывает на высокую избыточность входной информации, наличие специфических механизмов, уменьшающих количество информации по мере ее продвижения в структурах анализатора (от рецепторов к центральному отделу анализатора).
Из окружающей среды в организм в среднем поступает до 10 бит информации в секунду, но благодаря селективным свойствам сенсорных систем в мозг поступает лишь 10 бит информации. В процессе адаптивного приспособительного поведения животного организма значительная роль принадлежит сенсорным реле — промежуточным узловым структурам сенсорных систем. Они выполняют функции выявления во входных посылках физиологически важной информации. В результате в сенсорных реле, образующих фильтрующие (перекодирующие) центры, происходит регулирование суммарного входного информационного потока в соответствии с требованиями других отделов нервной системы и всего организма в целом.
Глава 4. НЕРВНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Спинной мозг
Морфофункциональная организация спинного мозга
Спинной мозг — наиболее древнее образование центральной нервной системы; он впервые появляется у ланцетника.
Приобретая новые связи и функции в ходе эволюции, спинной мозг высших организмов сохраняет старые связи и функции, которые у него возникли на всех предыдущих этапах развития.
Характерной чертой организации спинного мозга является периодичность его структуры в форме сегментов, имеющих входы в виде задних корешков, клеточную массу нейронов (серое вещество) и выходы в виде передних корешков (рис. 4.8).
Спинной мозг человека имеет 31—33 сегмента: 8 шейных (СI— CVIII), 12 грудных (ТI—TXII), 5 поясничных (LI—LV), S крестцовых (SI—SV), 1—3 копчиковых (CoI—СоIII).
Морфологических границ между сегментами спинного мозга не существует, поэтому деление на сегменты является функциональным и определяется зоной распределения в нем волокон заднего корешка и зоной клеток, которые образуют выход передних корешков. Каждый сегмент через свои корешки иннервирует три метамера тела и получает информацию также от трех метамеров тела. В итоге перекрытия каждый метамер тела иннервируется тремя сегментами и передает сигналы в три сегмента спинного мозга.
Спинной мозг человека имеет два утолщения: шейное и поясничное — в них содержится большее число нейронов, чем в остальных его участках.
Волокна, поступающие по задним корешкам спинного мозга, выполняют функции, которые определяются тем, где и на каких нейронах заканчиваются данные волокна.
В опытах с перерезкой и раздражением корешков спинного мозга показано, что задние корешки являются афферентными, чувствительными, центростремительными, а передние — эфферентными, двигательными, центробежными (закон Белла—Мажанди).
Афферентные входы в спинной мозг организованы аксонами спинальных ганглиев, лежащих вне спинного мозга, и аксонами экстра- и интрамуральных ганглиев симпатического и парасимпатического отделов автономной нервной системы.
Первая группа афферентных входов спинного мозга образована чувствительными волокнами, идущими от мышечных рецепторов, рецепторов сухожилий, надкостницы, оболочек суставов. Эта группа рецепторов образует начало так называемой проприоцептивной чувствительности. Проприоцептивные волокна по толщине и скорости проведения возбуждения делятся на 3 группы (табл. 4.1). Волокна каждой группы имеют свои пороги возникновения возбуждения.
Вторая группа афферентных входов спинного мозга начинается от кожных рецепторов: болевых, температурных, тактильных, давления — и представляет собой кожную рецептирующую систему.
Третья группа афферентных входов спинного мозга представлена рецептирующими входами от висцеральных органов; это висцеро-рецептивная система.
Эфферентные (двигательные) нейроны расположены в передних рогах спинного мозга, и их волокна иннервируют всю скелетную мускулатуру.
Особенности нейронной организации спинного мозга
Нейроны спинного мозга образуют его серое вещество в виде симметрично расположенных двух передних и двух задних рогов в шейном, поясничном и крестцовом отделах. Серое вещество распределено на ядра, вытянутые по длине спинного мозга, и на поперечном разрезе располагается в форме буквы Н. В грудном отделе спинной мозг имеет, помимо названных, еще и боковые рога (рис. 4.9).
Задние рога выполняют главным образом сенсорные функции и содержат нейроны, передающие сигналы в вышележащие центры, в симметричные структуры противоположной стороны либо к передним рогам спинного мозга.
В передних рогах находятся нейроны, дающие свои аксоны к мышцам. Все нисходящие пути центральной нервной системы, вызывающие двигательные реакции, заканчиваются на нейронах передних рогов. В связи с этим Шеррингтон назвал их «общим конечным путем».
Начиная с I грудного сегмента спинного мозга и до первых поясничных сегментов, в боковых рогах серого вещества располагаются нейроны симпатического, а в крестцовых — парасимпатического отдела автономной (вегетативной) нервной системы.
Спинной мозг человека содержит около 13 млн. нейронов, из них 3% — мотонейроны, а 97% — вставочные. Функционально нейроны спинного мозга можно разделить на 4 основные группы:
мотонейроны, или двигательные, — клетки передних рогов, аксоны которых образуют передние корешки;
интернейроны — нейроны, получающие информацию от спинальных ганглиев и располагающиеся в задних рогах. Эти нейроны реагируют на болевые, температурные, тактильные, вибрационные, проприоцептивные раздражения;
симпатические, парасимпатические нейроны расположены преимущественно в боковых рогах. Аксоны этих нейронов выходят из спинного мозга в составе передних корешков;
ассоциативные клетки — нейроны собственного аппарата спинного мозга, устанавливающие связи внутри и между сегментами.
В средней зоне серого вещества (между задним и передним рогами) спинного мозга имеется промежуточное ядро (ядро Кахаля) с клетками, аксоны которых идут вверх или вниз на 1—2 сегмента и дают коллатерали на нейроны ипси- и контралатеральной стороны, образуя сеть. Подобная сеть имеется и на верхушке заднего рога спинного мозга — эта сеть образует так называемое студенистое вещество (желатинозная субстанция Роланда) и выполняет функции ретикулярной формации спинного мозга.
Средняя часть серого вещества спинного мозга содержит преимущественно короткоаксонные веретенообразные клетки (промежуточные нейроны), выполняющие связующую функцию между симметричными отделами сегмента, между клетками его передних и задних рогов.
Мотонейроны. Аксон мотонейрона своими терминалами иннервирует сотни мышечных волокон, образуя мотонейронную единицу. Чем меньше мышечных волокон иннервирует один аксон (т. е. чем меньше количественно мотонейронная единица), тем более дифференцированные, точные движения выполняет мышца (см. раздел 2.4).
Несколько мотонейронов могут иннервировать одну мышцу, в этом случае они образуют так называемый мотонейронный пул. Возбудимость мотонейронов одного пула различна, поэтому при разной интенсивности раздражения в сокращение вовлекается разное количество волокон одной мышцы. При оптимальной силе раздражения сокращаются все волокна данной мышцы; в этом случае развивается максимальное сокращение мышцы.
Мотонейроны спинного мозга функционально делят на α- и γ-нейроны.
α-Мотонейроны образуют прямые связи с чувствительными путями, идущими от экстрафузальных волокон мышечного веретена, имеют до 20 000 синапсов на своих дендритах и характеризуются низкой частотой импульсации (10—20 в секунду), γ-Мотонейроны, иннервирующие интрафузальные мышечные волокна мышечного веретена, получают информацию о его состоянии через промежуточные нейроны. Сокращение интрафузального мышечного волокна не приводит к сокращению мышцы, но повышает частоту разрядов импульсов, идущих от рецепторов волокна в спинной мозг. Эти нейроны обладают высокой частотой импульсации (до 200 в секунду).
Интернейроны. Эти промежуточные нейроны, генерирующие импульсы с частотй до 1000 в секунду, являются фоновоактивными и имеют на своих дендритах до 500 синапсов. Функция интернейронов заключается в организации связей между структурами спинного мозга и обеспечении влияния восходящих и нисходящих путей на клетки отдельных сегментов спинного мозга. Очень важной функцией интернейронов является торможение активности нейронов, что обеспечивает сохранение направленности пути возбуждения. Возбуждение интернейронов, связанных с моторными клетками, оказывает тормозящее влияние на мышцы-антагонисты.
Нейроны симпатического отдела автономной системы. Расположены в боковых рогах сегментов грудного отдела спинного мозга. Эти нейроны являются фоновоактивными, но имеют редкую частоту импульсации (3—5 в секунду).
Нейроны парасимпатического отдела автономной системы. Локализуются в сакральном отделе спинного мозга и являются фоновоактивными.
В случаях раздражения и поражения задних корешков спинного мозга наблюдаются «стреляющие», опоясывающие боли на уровне метамера пораженного сегмента, снижение чувствительности всех видов, утрата или снижение рефлексов, вызываемых с метамера тела, который передает информацию в пораженный корешок.
В случаях изолированного поражения заднего рога утрачивается болевая и температурная чувствительность на стороне повреждения, а тактильная и проприоцептивная сохраняется, так как из заднего корешка аксоны температурной и болевой чувствительности идут в задний рог, а аксоны тактильной и проприоцептивной — прямо в задний столб и по проводящим путям поднимаются вверх.
Вследствие того, что аксоны вторых нейронов болевой и температурной чувствительности идут на противоположную сторону через переднюю серую спайку спинного мозга, при повреждении этой спайки на теле симметрично утрачивается болевая и температурная чувствительность.
Поражение переднего рога и переднего корешка спинного мозга приводит к параличу мышц, которые теряют тонус, атрофируются, при этом исчезают рефлексы, связанные с пораженным сегментом.
В случае поражения боковых рогов спинного мозга исчезают кожные сосудистые рефлексы, нарушается потоотделение, наблюдаются трофические изменения кожи, ногтей. При одностороннем поражении парасимпатического отдела автономной нервной системы на уровне крестцовых отделов спинного мозга нарушений дефекации и мочеиспускания не наблюдается, так как корковая иннервация этих центров является двусторонней.
Проводящие пути спинного мозга
Белое вещество спинного мозга состоит из миелиновых волокон, которые собраны в пучки. Эти волокна могут быть короткими (межсегментарные) и длинными — соединяющими разные отделы головного мозга со спинным и наоборот. Короткие волокна (их называют ассоциативными) связывают нейроны разных сегментов или симметричные нейроны противоположных сторон спинного мозга.
Длинные волокна (их называют проекционными) делятся на восходящие, идущие к головному мозгу, и нисходящие — идущие от головного мозга к спинному. Эти волокна образуют проводящие пути спинного мозга.
Пучки аксонов образуют вокруг серого вещества так называемые канатики: передние — расположенные кнутри от передних рогов, задние — расположенные между задними рогами серого вещества, и боковые — расположенные на латеральной стороне спинного мозга между передними и задними корешками.
Аксоны спинальных ганглиев и серого вещества спинного мозга идут в его белое вещество, а затем в другие структуры ЦНС, создавая тем самым восходящие и нисходящие проводящие пути.
В передних канатиках расположены нисходящие пути:
передний корково-спинномозговой, или пирамидный, путь (tractus corticospinalis ventralis, s.anterior), являющийся прямым неперекрещенным;
задний продольный пучок (fasciculus longitudinalis dorsalis, s.posterior);
покрышечно-спинномозговой, или тектоспинальный, путь (tractus tectospinalis);
преддверно-спинномозговой, или вестибулоспинальный, путь (tractus vestibulospinalis).
В задних канатиках проходят восходящие пути:
тонкий пучок, или пучок Голля (fasciculus gracilis);
клиновидный пучок, или пучок Бурдаха (fasciculus cuneatus).
В боковых канатиках проходят нисходящие и восходящие пути.
К нисходящим путям относятся:
латеральный корково-спинномозговой, или пирамидный, путь (tractus corticospinalis lateralis), является перекрещенным;
красноядерно-спинномозговой, или руброспинальный, путь (tractus rubrospinalis);
ретикулярно-спинномозговой, или ретикулоспинальный, путь (tractus reticulospinalis).
К восходящим путям относятся:
спинно-таламический (tractus spinothalamicus) путь;
латеральный и передний спинно-мозжечковые, или пучки Флексига и Говерса (tractus spinocerebellares lateralis et ventralis).
Ассоциативные, или проприоспинальные, пути связывают между собой нейроны одного или разных сегментов спинного мозга. Они начинаются от нейронов серого вещества промежуточной зоны, идут в белое вещество бокового или переднего канатиков спинного мозга и заканчиваются в сером веществе промежуточной зоны или на мотонейронах передних рогов других сегментов. Эти связи выполняют ассоциативную функцию, которая заключается в координации позы, тонуса мышц, движений разных метамеров туловища. К проприоспинальным путям относятся также комиссуральные волокна, соединяющие функционально однородные симметричные и несимметричные участки спинного мозга.
Нисходящие пути (рис. 4.10) связывают отделы головного мозга с моторными или вегетативными эфферентными нейронами.
Цереброспинальные нисходящие пути начинаются от нейронов структур головного мозга и заканчиваются на нейронах сегментов спинного мозга. Сюда относятся следующие пути: передний (прямой) и латеральный (перекрещенный) корково-спинномозговой (от пирамидных нейронов пирамидной и экстрапирамидной коры, обеспечивающие регуляцию произвольных движений), красноядерно-спинномозговой (руброспинальный), преддверно-спинномозговой (вестибулоспинальный), ретикулярно-спинномозговой (ретикулоспинальный) пути участвуют в регуляции тонуса мускулатуры. Объединяющим для всех перечисленных путей служит то, что конечным их пунктом являются мотонейроны передних рогов. У человека пирамидный путь оканчивается непосредственно на мотонейронах, а другие пути оканчиваются преимущественно на промежуточных нейронах.
Пирамидный путь состоит из двух пучков: латерального и прямого. Латеральный пучок начинается от нейронов коры большого мозга, на уровне продолговатого мозга переходит на другую сторону, образуя перекрест, и спускается по противоположной стороне спинного мозга. Прямой пучок спускается до своего сегмента и там переходит к мотонейронам противоположной стороны. Следовательно, весь пирамидный путь является перекрещенным.
Красноядерно-спинномозговой, или руброспинальный, путь (tractus rubrospinalis) состоит из аксонов нейронов красного ядра. Эти аксоны сразу после выхода из ядра переходят на симметричную сторону и делятся на три пучка. Один идет в спинной мозг, другой в мозжечок, третий — в ретикулярную формацию ствола мозга.
Нейроны, дающие начало этому пути, участвуют в управлении мышечным тонусом. Рубромозжечковый и руброретикулярные пути обеспечивают координацию активности пирамидных нейронов коры и нейронов мозжечка, участвующих в организации произвольных движений.
Преддверно-спинномозговой, или вестибулоспинальный, путь (tractus vestibulospinalis) начинается от нейронов латерального преддверного ядра (ядра Дейтерса), лежащего в продолговатом мозге. Это ядро регулирует активность мотонейронов спинного мозга, обеспечивает тонус мускулатуры, согласованность движений, равновесие.
Ретикулярно-спинномозговой, или ретикулоспинальный, путь (tractus reticulospinalis) идет от ретикулярной формации ствола мозга к мотонейронам спинного мозга, через него ретикулярная формация регулирует тонус мускулатуры.
Повреждения проводникового аппарата спинного мозга приводят к нарушениям двигательной или чувствительной системы ниже участка повреждения.
Пересечение пирамидального пути вызывает ниже перерезки гипертонус мышц (мотонейроны спинного мозга освобождаются от тормозного влияния пирамидных клеток коры) и, как следствие, к спастическому параличу.
При пересечении чувствительных путей полностью утрачивается мышечная, суставная, болевая и другая чувствительность ниже места перерезки спинного мозга.
Спиноцеребральные восходящие пути (см. рис. 4.10) соединяют сегменты спинного мозга со структурами головного мозга. Эти пути представлены путями проприоцептивной чувствительности, таламическим, спинно-мозжечковым, спинно-ретикулярным. Их функция заключается в передаче информации в мозг об экстеро-, интеро- и проприорецептивных раздражениях.
Проприоцептивный путь (тонкий и клиновидный пучки) начинается от рецепторов глубокой чувствительности мышц сухожилий, надкостницы, оболочек суставов. Тонкий пучок начинается от ганглиев, собирающих информацию от каудальных отделов тела, таза, нижних конечностей. Клиновидный пучок начинается от ганглиев, собирающих информацию от мышц грудной клетки, верхних конечностей. От спинального ганглия аксоны идут в задние корешки спинного мозга, в белое вещество задних канатиков, поднимаются в тонкое и клиновидные ядра продолговатого мозга. Здесь происходит первое переключение на новый нейрон, далее путь идет в латеральные ядра таламуса противоположного полушария большого мозга, переключается на новый нейрон, т. е. происходит второе переключение. От таламуса путь поднимается к нейронам IV слоя соматосенсорной области коры. Волокна этих трактов отдают коллатерали в каждом сегменте спинного мозга, что создает возможность коррекции позы всего туловища. Скорость проведения возбуждения по волокнам данного тракта достигает 60—100 м/с.
Спинно-таламический путь (tractus spinothalamicus) — основной путь кожной чувствительности — начинается от болевых, температурных, тактильных рецепторов и барорецепторов кожи. Болевые, температурные, тактильные сигналы от рецепторов кожи идут в спинальный ганглий, далее через задний корешок к заднему рогу спинного мозга (первое переключение). Чувствительные нейроны задних рогов посылают аксоны на противоположную сторону спинного мозга и поднимаются по боковому канатику к таламусу; скорость проведения возбуждения по ним 1—30 м/с (второе переключение), отсюда — в сенсорную область коры большого мозга. Часть волокон кожных рецепторов идет к таламусу по переднему канатику спинного мозга.
Спинно-мозжечковые пути (tractus spinocerebellares) лежат в боковых канатиках спинного мозга и представлены неперекрещивающимися передним, спинно-мозжечковым путем (пучок Говерса) и дважды перекрещивающимся задним спинно-мозжечковым путем (пучок Флексига). Следовательно, все спинно-мозжечковые пути начинаются на левой стороне тела и заканчиваются в левой доле мозжечка; точно также и правая доля мозжечка получает информацию только со своей стороны тела. Эта информация идет от сухожильных рецепторов Гольджи, проприорецепторов, рецепторов давления, прикосновения. Скорость проведения возбуждения по этим трактам достигает 110—120 м/с.
Рефлекторные функции спинного мозга
Функциональное разнообразие нейронов спинного мозга, наличие в нем афферентных нейронов, интернейронов, мотонейронов и нейронов автономной нервной системы, а также многочисленных прямых и обратных, сегментарных, межсегментарных связей и связей со структурами головного мозга — все это создает условия для рефлекторной деятельности спинного мозга с участием как собственных структур, так и головного мозга. Подобная организация позволяет реализовывать все двигательные рефлексы тела, диафрагмы, мочеполовой системы и прямой кишки, терморегуляции, сосудистые рефлексы и т. д.
Рефлекторные реакции спинного мозга зависят от места, силы раздражения, площади раздражаемой рефлексогенной зоны, скорости проведения по афферентным и эфферентным волокнам и, наконец, от влияния головного мозга. Сила и длительность рефлексов спинного мозга увеличивается при повторении раздражения (суммация).
Собственная рефлекторная деятельность спинного мозга осуществляется сегментарными рефлекторными дугами.
Сегментарная рефлекторная дуга состоит из рецептивного поля, из которого импульсация по чувствительному волокну нейрона спинального ганглия, а затем по аксону этого же нейрона через задний корешок входит в спинной мозг, далее аксон может идти прямо к мотонейрону переднего рога, аксон которого подходит к мышце. Так образуется моносинаптическая рефлекторная дуга, которая имеет один синапс между афферентным нейроном спинального ганглия и мотонейроном переднего рога. Эти рефлекторные дуги образуются в таких рефлексах, которые возникают только при раздражении рецепторов аннулоспиральных окончаний мышечных веретен.
Другие спинальные рефлексы реализуются с участием интернейронов заднего рога или промежуточной области спинного мозга. В итоге возникают полисинаптические рефлекторные дуги.
Рассмотрим некоторые рефлексы спинного мозга
Миотатические рефлексы — рефлексы на растяжение мышцы. Быстрое растяжение мышцы, всего на несколько миллиметров механическим ударом по ее сухожилию приводит к сокращению всей мышцы и двигательной реакции. Например, легкий удар по сухожилию надколенной чашечки вызывает сокращение мышц бедра и разгибание голени. Дуга этого рефлекса следующая: мышечные рецепторы четырехглавой мышцы бедра à спинальный ганглий à задние корешки à задние рога III поясничного сегмента à мотонейроны передних рогов того же сегмента à экстрафузальные волокна четырехглавой мышцы бедра. Реализация этого рефлекса была бы невозможна, если бы одновременно с сокращением мышц-разгибателей не расслаблялись мышцы-сгибатели. Рефлекс на растяжение свойствен всем мышцам, но у мышц-разгибателей, они хорошо выражены и легко вызываются.
Рефлексы с рецепторов кожи носят характер, зависящий от силы раздражения, вида раздражаемого рецептора, но чаще всего конечная реакция выглядит в виде усиления сокращения мышц-сгибателей.
Висцеромоторные рефлексы возникают при стимуляции афферентных нервов внутренних органов и характеризуются появлением двигательных реакций мышц грудной клетки и брюшной стенки, мышц разгибателей спины.
Рефлексы автономной нервной системы имеют свои пути. Они начинаются от различных рецепторов, входят в спинной мозг через задние корешки, задние рога, далее в боковые рога, нейроны которых через передний корешок посылают аксоны не непосредственно к органам, а к ганглию симпатического или парасимпатического отдела автономной нервной системы.
Автономные (вегетативные) рефлексы обеспечивают реакцию внутренних органов, сосудистой системы на раздражение висцеральных, мышечных, кожных рецепторов. Эти рефлексы отличаются большим латентным периодом (ЛП) двумя фазами реакции: первая — ранняя — возникает с ЛП 7—9 мс и реализуется ограниченным числом сегментов, вторая — поздняя — возникает с большим ЛП — до 21 мс и вовлекает в реакцию практически все сегменты спинного мозга. Поздний компонент вегетативного рефлекса обусловлен вовлечением в него вегетативных центров головного мозга.
Сложной формой рефлекторной деятельности спинного мозга является рефлекс, реализующий произвольное движение. В основе реализации произвольного движения лежит γ-афферентная рефлекторная система. В нее входят пирамидная кора, экстрапирамидная система, α- и γ-мотонейроны спинного мозга, экстра- и интрафузальные волокна мышечного веретена.
При травмах у человека в ряде случаев происходит полное или половинное пересечение спинного мозга. При половинном латеральном повреждении спинного мозга развивается синдром Броун-Секара. Он проявляется в том, что на стороне поражения спинного мозга (ниже места поражения) развивается паралич двигательной системы вследствие повреждения пирамидных путей. На противоположной поражению стороне движения сохраняются.
На стороне поражения (ниже места поражения) нарушается проприоцептивная чувствительность. Это обусловлено тем, что восходящие пути глубокой чувствительности идут по своей стороне спинного мозга до продолговатого мозга, где происходит их перекрест.
На противоположной стороне туловища (относительно повреждения спинного мозга) нарушается болевая чувствительность, так как проводящие пути болевой чувствительности кожи идут от спинального ганглия в задний рог спинного мозга, где переключаются на новый нейрон, аксон которого переходит на противоположную сторону. В итоге если повреждена левая половина спинного мозга, то исчезает болевая чувствительность правой половины туловища ниже повреждения. Полную перерезку спинного мозга в экспериментах на животных производят для исследования влияния вышележащих отделов ЦНС на нижележащие. После полного пересечения спинного мозга возникает спинальный шок. Это явление заключается в том, что все центры ниже перерезки перестают организовывать присущие им рефлексы. Нарушение рефлекторной деятельности после пересечения спинного мозга у разных животных длится разное время. У лягушек оно исчисляется десятками секунд, у кролика рефлексы восстанавливаются через 10—15 мин, у собак отдельные рефлексы, например, мышечного сокращения, восстанавливаются через несколько часов, другие — через несколько дней (рефлексы регуляции артериального давления), через недели восстанавливаются рефлексы мочеиспускания. У обезьян первые признаки восстановления рефлексов после перерезки спинного мозга появляются через несколько суток; у человека первые спинальные рефлексы восстанавливаются через несколько недель, а то и месяцев.
Следовательно, чем сложнее организация ЦНС у животного, тем сильнее контроль вышележащих отделов мозга над нижележащими. То, что причиной шока является нарушение супраспинальных влияний, доказывается повторной перерезкой спинного мозга ниже места первой перерезки. В этом случае спинальный шок не возникает, рефлекторная деятельность спинного мозга сохраняется.
По истечении длительного периода времени после шока спинальные рефлексы резко усиливаются, что объясняется устранением тормозного влияния ретикулярной формации ствола мозга на рефлексы спинного мозга.
Ствол мозга
Ствол мозга включает продолговатый мозг, мост, средний мозг, промежуточный мозг и мозжечок. Ствол мозга выполняет следующие функции:
1) организует рефлексы, обеспечивающие подготовку и реализацию различных форм поведения; 2)осуществляет проводниковую функцию: через ствол мозга проходят в восходящем и нисходящем направлении пути, связывающие между собой структуры ЦНС; 3) при организации поведения обеспечивает взаимодействие своих структур между собой, со спинным мозгом, базальными ганглиями и корой большого мозга, т. е. обеспечивает ассоциативную функцию.
Продолговатый мозг
Особенности функциональной организации. Продолговатый мозг (medulla oblongata) у человека имеет длину около 25 мм. Он является продолжением спинного мозга. Структурно по разнообразию и строению ядер продолговатый мозг сложнее, чем спинной. В отличие от спинного мозга он не имеет метамерного, повторяемого строения, серое вещество в нем расположено не в центре, а ядрами к периферии.
В продолговатом мозге находятся оливы, связанные со спинным мозгом, экстрапирамидной системой и мозжечком — это тонкое и клиновидное ядра проприоцептивной чувствительности (ядра Голля и Бурдаха). Здесь же находятся перекресты нисходящих пирамидных путей и восходящих путей, образованных тонким и клиновидным пучками (Голля и Бурдаха), ретикулярная формация.
Продолговатый мозг за счет своих ядерных образований и ретикулярной формации участвует в реализации вегетативных, соматических, вкусовых, слуховых, вестибулярных рефлексов. Особенностью продолговатого мозга является то, что его ядра, возбуждаясь последовательно, обеспечивают выполнение сложных рефлексов, требующих последовательного включения разных мышечных групп, что наблюдается, например, при глотании.
В продолговатом мозге расположены ядра следующих черепных нервов:
пара VIII черепных нервов — преддверно-улитковый нерв состоит из улитковой и преддверной частей. Улитковое ядро лежит в продолговатом мозге;
пара IX — языкоглоточный нерв (п. glossopharyngeus); его ядро образовано 3 частями — двигательной, чувствительной и вегетативной. Двигательная часть участвует в иннервации мышц глотки и полости рта, чувствительная — получает информацию от рецепторов вкуса задней трети языка; вегетативная иннервирует слюнные железы;
пара X — блуждающий нерв (n.vagus) имеет 3 ядра: вегетативное иннервирует гортань, пищевод, сердце, желудок, кишечник, пищеварительные железы; чувствительное получает информацию от рецепторов альвеол легких и других внутренних органов и двигательное (так называемое обоюдное) обеспечивает последовательность сокращения мышц глотки, гортани при глотании;
пара XI — добавочный нерв (n.accessorius); его ядро частично расположено в продолговатом мозге;
пара XII — подъязычный нерв (n.hypoglossus) является двигательным нервом языка, его ядро большей частью расположено в продолговатом мозге.
Сенсорные функции. Продолговатый мозг регулирует ряд сенсорных функций: рецепцию кожной чувствительности лица — в сенсорном ядре тройничного нерва; первичный анализ рецепции вкуса — в ядре языкоглоточного нерва; рецепцию слуховых раздражений — в ядре улиткового нерва; рецепцию вестибулярных раздражений — в верхнем вестибулярном ядре. В задневерхних отделах продолговатого мозга проходят пути кожной, глубокой, висцеральной чувствительности, часть из которых переключается здесь на второй нейрон (тонкое и клиновидное ядра). На уровне продолготоватого мозга перечисленные сенсорные функции реализуют первичный анализ силы и качества раздражения, далее обработанная информация передается в подкорковые структуры для определения биологической значимости данного раздражения.
Проводниковые функции. Через продолготоватый мозг проходят все восходящие и нисходящие пути спинного мозга: спинно-таламический, кортикоспинальный, руброспинальный. В нем берут начало вестибулоспинальный, оливоспинальный и ретикулоспинальный тракты, обеспечивающие тонус и координацию мышечных реакций. В продолговатом мозге заканчиваются пути из коры большого мозга — корковоретикулярные пути. Здесь заканчиваются восходящие пути проприоцептивной чувствительности из спинного мозга: тонкого и клиновидного. Такие образования головного мозга, как мост, средний мозг, мозжечок, таламус, гипоталамус и кора большого мозга, имеют двусторонние связи с продолговатым мозгом. Наличие этих связей свидетельствует об участии продолговатого мозга в регуляции тонуса скелетной мускулатуры, вегетативных и высших интегративных функций, анализе сенсорных раздражений.
Рефлекторные функции. Многочисленные рефлексы продолговатого мозга делят на жизненно важные и нежизненно важные, однако такое представление достаточно условно. Дыхательные и сосудодвигательные центры продолговатого мозга можно отнести к жизненно важным центрам, так как в них замыкается ряд сердечных и дыхательных рефлексов.
Продолговатый мозг организует и реализует ряд защитных рефлексов: рвоты, чиханья, кашля, слезоотделения, смыкания век. Эти рефлексы реализуются благодаря тому, что информация о раздражении рецепторов слизистой оболочки глаза, полости рта, гортани, носоглотки через чувствительные ветви тройничного и языкоглоточного нервов попадает в ядра продолговатого мозга, отсюда идет команда к двигательным ядрам тройничного, блуждающего, лицевого, языкоглоточного, добавочного или подъязычного нервов, в результате реализуется тот или иной защитный рефлекс. Точно так же за счет последовательного включения мышечных групп головы, шеи, грудной клетки и диафрагмы организуются рефлексы пищевого поведения: сосания, жевания, глотания.
Кроме того, продолговатый мозг организует рефлексы поддержания позы. Эти рефлексы формируются за счет афферентации от рецепторов преддверия улитки и полукружных каналов в верхнее вестибулярное ядро; отсюда переработанная информация оценки необходимости изменения позы посылается к латеральному и медиальному вестибулярным ядрам. Эти ядра участвуют в определении того, какие мышечные системы, сегменты спинного мозга должны принять участие в изменении позы, поэтому от нейронов медиального и латерального ядра по вестибулоспинальному пути сигнал поступает к передним рогам соответствующих сегментов спинного мозга, иннервирующих мышцы, участие которых в изменении позы в данный момент необходимо.
Изменение позы осуществляется за счет статических и статокинетических рефлексов. Статические рефлексы регулируют тонус скелетных мышц с целью удержания определенного положения тела. Статокинетические рефлексы продолговатого мозга обеспечивают перераспределение тонуса мышц туловища для организации позы, соответствующей моменту прямолинейного или вращательного движения.
Большая часть автономных рефлексов продолговатого мозга реализуется через расположенные в нем ядра блуждающего нерва, которые получают информацию о состоянии деятельности сердца, сосудов, пищеварительного тракта, легких, пищеварительных желез и др. В ответ на эту информацию ядра организуют двигательную и секреторную реакции названных органов.
Возбуждение ядер блуждающего нерва вызывает усиление сокращения гладких мышц желудка, кишечника, желчного пузыря и одновременно расслабление сфинктеров этих органов. При этом замедляется и ослабляется работа сердца, сужается просвет бронхов.
Деятельность ядер блуждающего нерва проявляется также в усилении секреции бронхиальных, желудочных, кишечных желез, в возбуждении поджелудочной железы, секреторных клеток печени.
В продолговатом мозге локализуется центр слюноотделения, парасимпатическая часть которого обеспечивает усиление общей секреции, а симпатическая — белковой секреции слюнных желез.
В структуре ретикулярной формации продолговатого мозга расположены дыхательный и сосудодвигательный центры. Особенность этих центров в том, что их нейроны способны возбуждаться рефлекторно и под действием химических раздражителей.
Дыхательный центр локализуется в медиальной части ретикулярной формации каждой симметричной половины продолговатого мозга и разделен на две части, вдоха и выдоха.
В ретикулярной формации продолговатого мозга представлен другой жизненно важный центр — сосудодвигательный центр (регуляции сосудистого тонуса). Он функционирует совместно с вышележащими структурами мозга и прежде всего с гипоталамусом. Возбуждение сосудодвигательного центра всегда изменяет ритм дыхания, тонус бронхов, мышц кишечника, мочевого пузыря, цилиарной мышцы и др. Это обусловлено тем, что ретикулярная формация продолговатого мозга имеет синаптические связи с гипоталамусом и другими центрами.
В средних отделах ретикулярной формации находятся нейроны, образующие ретикулоспинальный путь, оказывающий тормозное влияние на мотонейроны спинного мозга. На дне IV желудочка расположены нейроны «голубого пятна». Их медиатором является норадреналин. Эти нейроны вызывают активацию ретикулоспинального пути в фазу «быстрого» сна, что приводит к торможению спинальных рефлексов и снижению мышечного тонуса.
Симптомы повреждения. Повреждение левой или правой половины продолговато мозга выше перекреста восходящих путей проприоцептивной чувствительности вызывает на стороне повреждения нарушения чувствительности и работы мышц лица и головы. В то же время на противоположной стороне относительно стороны повреждения наблюдаются нарушения кожной чувствительности и двигательные параличи туловища и конечностей. Это объясняется тем, что восходящие и нисходящие проводящие пути из спинного мозга и в спинной мозг перекрещиваются, а ядра черепных нервов иннервируют свою половину головы, т. е. черепные нервы не перекрещиваются.
Мост
Мост (pons cerebri, pons Varolii) располагается выше продолговатого мозга и выполняет сенсорные, проводниковые, двигательные, интегративные рефлекторные функции.
В состав моста входят ядра лицевого, тройничного, отводящего, преддверно-улиткового нерва (вестибулярные и улитковые ядра), ядра преддверной части преддверно-улиткового нерва (вестибулярного нерва): латеральное (Дейтерса) и верхнее (Бехтерева). Ретикулярная формация моста тесно связана с ретикулярной формацией среднего и продолговатого мозга.
Важной структурой моста является средняя ножка мозжечка. Именно она обеспечивает функциональные компенсаторные и морфологические связи коры большого мозга с полушариями мозжечка.
Сенсорные функции моста обеспечиваются ядрами преддверно-улиткового, тройничного нервов. Улитковая часть преддверно-улиткового нерва заканчивается в мозге в улитковых ядрах; преддверная часть преддверно-улиткового нерва — в треугольном ядре, ядре Дейтерса, ядре Бехтерева. Здесь происходит первичный анализ вестибулярных раздражений их силы и направленности.
Чувствительное ядро тройничного нерва получает сигналы от рецепторов кожи лица, передних отделов волосистой части головы, слизистой оболочки носа и рта, зубов и конъюнктивы глазного яблока. Лицевой нерв (п. facialis) иннервирует все мимические мышцы лица. Отводящий нерв (п. abducens) иннервирует прямую латеральную мышцу, отводящую глазное яблоко кнаружи.
Двигательная порция ядра тройничного нерва (п. trigeminus) иннервирует жевательные мышцы, мышцу, натягивающую барабанную перепонку, и мышцу, натягивающую небную занавеску.
Проводящая функция моста. Обеспечивается продольно и поперечно расположенными волокнами. Поперечно расположенные волокна образуют верхний и нижний слои, а между ними проходят идущие из коры большого мозга пирамидные пути. Между поперечными волокнами расположены нейронные скопления — ядра моста. От их нейронов начинаются поперечные волокна, которые идут на противоположную сторону моста, образуя среднюю ножку мозжечка и заканчиваясь в его коре.
В покрышке моста располагаются продольно идущие пучки волокон медиальной петли (lemniscus medialis). Они пересекаются поперечно идущими волокнами трапециевидного тела (corpus trapezoideum), представляющие собой аксоны улитковой части преддверно-улиткового нерва противоположной стороны, которые заканчиваются в ядре верхней оливы (oliva superior). От этого ядра идут пути боковой петли (lemniscus lateralis), которые направляются в заднее четверохолмие среднего мозга и в медиальные коленчатые тела промежуточного мозга.
В покрышке мозга локализуются переднее и заднее ядра трапециевидного тела и латеральной петли. Эти ядра вместе с верхней оливой обеспечивают первичный анализ информации от органа слуха и затем передают информацию в задние бугры четверохолмий.
В покрышке также расположены длинный медиальный и тектоспинальный пути.
Собственные нейроны структуры моста образуют его ретикулярную формацию, ядра лицевого, отводящего нервов, двигательной порции ядра и среднее сенсорное ядро тройничного нерва.
Ретикулярная формация моста является продолжением ретикулярной формации продолговатого мозга и началом этой же системы среднего мозга. Аксоны нейронов ретикулярной формации моста идут в мозжечок, в спинной мозг (ретикулоспинальный путь). Последние активируют нейроны спинного мозга.
Ретикулярная формация моста влияет на кору большого мозга, вызывая ее пробуждение или сонное состояние. В ретикулярной формации моста находятся две группы ядер, которые относятся к общему дыхательному центру. Один центр активирует центр вдоха продолговатого мозга, другой — центр выдоха. Нейроны дыхательного центра, расположенные в мосте, адаптируют работу дыхательных клеток продолговатого мозга в соответствии с меняющимся состоянием организма.
Средний мозг
Морфофункциональная организация. Средний мозг (mesencephalon) представлен четверохолмием и ножками мозга. Наиболее крупными ядрами среднего мозга являются красное ядро, черное вещество и ядра черепных (глазодвигательного и блокового) нервов, а также ядра ретикулярной формации.
Сенсорные функции. Реализуются за счет поступления в него зрительной, слуховой информации.
Проводниковая функция. Заключается в том, что через него проходят все восходящие пути к вышележащим таламусу (медиальная петля, спииноталамический путь), большому мозгу и мозжечку. Нисходящие пути идут через средний мозг к продолговатому и спинному мозгу. Это пирамидный путь, корково-мостовые волокна, руброретикулоспинальный путь.
Двигательная функция. Реализуется за счет ядра блокового нерва (n. trochlearis), ядер глазодвигательного нерва (п. oculomotorius), красного ядра (nucleus ruber), черного вещества (substantia nigra).
Красные ядра располагаются в верхней части ножек мозга. Они связаны с корой большого мозга (нисходящие от коры пути), подкорковыми ядрами, мозжечком, спинным мозгом (красноядерно-спинномозговой путь). Базальные ганглии головного мозга, мозжечок имеют свои окончания в красных ядрах. Нарушение связей красных ядер с ретикулярной формацией продолговатого мозга ведет к децеребрационной ригидности. Это состояние характеризуется сильным напряжением мышц-разгибателей конечностей, шеи, спины. Основной причиной возникновения децеребрационной ригидности служит выраженное активирующее влияние латерального вестибулярного ядра (ядро Дейтерса) на мотонейроны разгибателей. Это влияние максимально в отсутствие тормозных влияний красного ядра и вышележащих структур, а также мозжечка. При перерезке мозга ниже ядра латерального вестибулярного нерва децеребрационная ригидность исчезает.
Красные ядра, получая информацию от двигательной зоны коры большого мозга, подкорковых ядер и мозжечка о готовящемся движении и состоянии опорно-двигательного аппарата, посылают корригирующие импульсы к мотонейронам спинного мозга по руброспинальному тракту и тем самым регулируют тонус мускулатуры, подготавливая его уровень к намечающемуся произвольному движению.
Другое функционально важное ядро среднего мозга — черное вещество — располагается в ножках мозга, регулирует акты жевания, глотания (их последовательность), обеспечивает точные движения пальцев кисти руки, например при письме. Нейроны этого ядра способны синтезировать медиатор дофамин, который поставляется аксональным транспортом к базальным ганглиям головного мозга. Поражение черного вещества приводит к нарушению пластического тонуса мышц. Тонкая регуляция пластического тонуса при игре на скрипке, письме, выполнении графических работ обеспечивается черным веществом. В то же время при длительном удержании определенной позы происходят пластические изменения в мышцах за счет изменения их коллоидных свойств, что обеспечивает наименьшие затраты энергии. Регуляция этого процесса осуществляется клетками черного вещества.
Нейроны ядер глазодвигательного и блокового нервов регулируют движение глаза вверх, вниз, наружу, к носу и вниз к углу носа. Нейроны добавочного ядра глазодвигательного нерва (ядро Якубовича) регулируют просвет зрачка и кривизну хрусталика.
Рефлекторные функции. Функционально самостоятельными структурами среднего мозга являются бугры четверохолмия. Верхние из них являются первичными подкорковыми центрами зрительного анализатора (вместе с латеральными коленчатыми телами промежуточного мозга), нижние — слухового (вместе с медиальными коленчатыми телами промежуточного мозга). В них происходит первичное переключение зрительной и слуховой информации. От бугров четверохолмия аксоны их нейронов идут к ретикулярной формации ствола, мотонейронам спинного мозга. Нейроны четверохолмия могут быть полимодальными и детекторными. В последнем случае они реагируют только на один признак раздражения, например смену света и темноты, направление движения светового источника и т. д. Основная функция бугров четверохолмия — организация реакции настораживания и так называемых старт-рефлексов на внезапные, еще не распознанные, зрительные или звуковые сигналы. Активация среднего мозга в этих случаях через гипоталамус приводит к повышению тонуса мышц, учащению сокращений сердца; происходит подготовка к избеганию, к оборонительной реакции.
Четверохолмие организует ориентировочные зрительные и слуховые рефлексы.
У человека четверохолмный рефлекс является сторожевым. В случаях повышенной возбудимости четверохолмий при внезапном звуковом или световом раздражении у человека возникает вздрагивание, иногда вскакивание на ноги, вскрикивание, максимально быстрое удаление от раздражителя, подчас безудержное бегство.
При нарушении четверохолмного рефлекса человек не может быстро переключаться с одного вида движения на другое. Следовательно, четверохолмия принимают участие в организации произвольных движений.
Ретикулярная формация ствола мозга
Ретикулярная формация (formatio reticularis; РФ) мозга представлена сетью нейронов с многочисленными диффузными связями между собой и практически со всеми структурами центральной нервной системы. РФ располагается в толще серого вещества продолговатого, среднего, промежуточного мозга и изначально связана с РФ спинного мозга. В связи с этим целесообразно ее рассмотреть как единую систему. Сетевые связи нейронов РФ между собой позволили Дейтерсу назвать ее ретикулярной формацией мозга.
РФ имеет прямые и обратные связи с корой большого мозга, базальными ганглиями, промежуточным мозгом, мозжечком, средним, продолговатым и спинным мозгом.
Основной функцией РФ является регуляция уровня активности коры большого мозга, мозжечка, таламуса, спинного мозга.
С одной стороны, генерализованный характер влияния РФ на многие структуры мозга дал основание считать ее неспецифической системой. Однако исследования с раздражением РФ ствола показали, что она может избирательно оказывать активирующее или тормозящее влияние на разные формы поведения, на сенсорные, моторные, висцеральные системы мозга. Сетевое строение обеспечивает высокую надежность функционирования РФ, устойчивость к повреждающим воздействиям, так как локальные повреждения всегда компенсируются за счет сохранившихся элементов сети. С другой стороны, высокая надежность функционирования РФ обеспечивается тем, что раздражение любой из ее частей отражается на активности всей РФ данной структуры за счет диффузности связей.
Большинство нейронов РФ имеет длинные дендриты и короткий аксон. Существуют гигантские нейроны с длинным аксоном, образующие пути из РФ в другие области мозга, например в нисходящем направлении, ретикулоспинальный и руброспинальный. Аксоны нейронов РФ образуют большое число коллатералей и синапсов, которые оканчиваются на нейронах различных отделов мозга. Аксоны нейронов РФ, идущие в кору большого мозга, заканчиваются здесь на дендритах I и II слоев.
Активность нейронов РФ различна и в принципе сходна с активностью нейронов других структур мозга, но среди нейронов РФ имеются такие, которые обладают устойчивой ритмической активностью, не зависящей от приходящих сигналов.
В то же время в РФ среднего мозга и моста имеются нейроны, которые в покое «молчат», т. е. не генерируют импульсы, но возбуждаются при стимуляции зрительных или слуховых рецепторов. Это так называемые специфические нейроны, обеспечивающие быструю реакцию на внезапные, неопознанные сигналы. Значительное число нейронов РФ являются полисенсорными.
В РФ продолговатого, среднего мозга и моста конвергируют сигналы различной сенсорности. На нейроны моста приходят сигналы преимущественно от соматосенсорных систем. Сигналы от зрительной и слуховой сенсорных систем в основном приходят на нейроны РФ среднего мозга.
РФ контролирует передачу сенсорной информации, идущей через ядра таламуса, за счет того, что при интенсивном внешнем раздражении нейроны неспецифических ядер таламуса затормаживаются, тем самым снимается их тормозящее влияние с релейных ядер того же таламуса и облегчается передача сенсорной информации в кору большого мозга.
В РФ моста, продолговатого, среднего мозга имеются нейроны, которые реагируют на болевые раздражения, идущие от мышц или внутренних органов, что создает общее диффузное дискомфортное, не всегда четко локализуемое, болевое ощущение «тупой боли».
Повторение любого вида стимуляции приводит к снижению импульсной активности нейронов РФ, т. е. процессы адаптации (привыкания) присущи и нейронам РФ ствола мозга.
РФ ствола мозга имеет прямое отношение к регуляции мышечного тонуса, поскольку на РФ ствола мозга поступают сигналы от зрительного и вестибулярного анализаторов и мозжечка. От РФ к мотонейронам спинного мозга и ядер черепных нервов поступают сигналы, организующие положение головы, туловища и т. д.
Ретикулярные пути, облегчающие активность моторных систем спинного мозга, берут начало от всех отделов РФ. Пути, идущие от моста, тормозят активность мотонейронов спинного мозга, иннервирующих мышцы-сгибатели, и активируют мотонейроны мышц-разгибателей. Пути, идущие от РФ продолговатого мозга, вызывают противоположные эффекты. Раздражение РФ приводит к тремору, повышению тонуса мышц. После прекращения раздражения вызванный им эффект сохраняется длительно, видимо, за счет циркуляции возбуждения в сети нейронов.
РФ ствола мозга участвует в передаче информации от коры большого мозга, спинного мозга к мозжечку и, наоборот, от мозжечка к этим же системам. Функция данных связей заключается в подготовке и реализации моторики, связанной с привыканием, ориентировочными реакциями, болевыми реакциями, организацией ходьбы, движениями глаз.
Регуляция вегетативной деятельности организма РФ описана в разделе 4.3, здесь же заметим, что наиболее четко эта регуляция проявляется в функционировании дыхательного и сердечно-сосудистых центров. В регуляции вегетативных функций большое значение имеют так называемые стартовые нейроны РФ. Они дают начало циркуляции возбуждения внутри группы нейронов, обеспечивая тонус регулируемых вегетативных систем.
Влияния РФ можно разделить в целом на нисходящие и восходящие. В свою очередь каждое из этих влияний имеет тормозное и возбуждающее действие.
Восходящие влияния РФ на кору большого мозга повышают ее тонус, регулируют возбудимость ее нейронов, не изменяя специфику ответов на адекватные раздражения. РФ влияет на функциональное состояние всех сенсорных областей мозга, следовательно, она имеет значение в интеграции сенсорной информации от разных анализаторов.
РФ имеет прямое отношение к регуляции цикла бодрствование—сон. Стимуляция одних структур РФ приводит к развитию сна, стимуляция других вызывает пробуждение. Г. Мэгун и Д. Моруцци выдвинули концепцию, согласно которой все виды сигналов, идущих от периферических рецепторов, достигают по коллатералям РФ продолговатого мозга и моста, где переключаются на нейроны, дающие восходящие пути в таламус и затем в кору большого мозга.
Возбуждение РФ продолговатого мозга или моста вызывает синхронизацию активности коры большого мозга, появление медленных ритмов в ее электрических показателях, сонное торможение.
Возбуждение РФ среднего мозга вызывает противоположный эффект пробуждения: десинхронизацию электрической активности коры, появление быстрых низкоамплитудных β-подобных ритмов в электроэнцефалограмме.
Г. Бремер (1935) показал, что если перерезать мозг между передними и задними буграми четверохолмия, то животное перестает реагировать на все виды сигналов; если же перерезку произвести между продолговатым и средним мозгом (при этом РФ сохраняет связь с передним мозгом), то животное реагирует на свет, звук и другие сигналы. Следовательно, поддержание активного анализирующего состояния мозга возможно при сохранении связи с передним мозгом.
Реакция активации коры большого мозга наблюдается при раздражении РФ продолговатого, среднего, промежуточного мозга. В то же время раздражение некоторых ядер таламуса приводит к возникновению ограниченных локальных участков возбуждения, а не к общему ее возбуждению, как это бывает при раздражении других отделов РФ.
РФ ствола мозга может оказывать не только возбуждающее, но и тормозное влияние на активность коры мозга.
Нисходящие влияния РФ ствола мозга на регуляторную деятельность спинного мозга были установлены еще И. М. Сеченовым (1862). Им было показано, что при раздражении среднего мозга кристалликами соли у лягушки рефлексы отдергивания лапки возникают медленно, требуют более сильного раздражения или не появляются вообще, т. е. тормозятся.
Г. Мэгун (1945—1950), нанося локальные раздражения на РФ продолговатого мозга, нашел, что при раздражении одних точек тормозятся, становятся вялыми рефлексы сгибания передней лапы, коленный, роговичный. При раздражении РФ в других точках продолговатого мозга эти же рефлексы вызывались легче, были сильнее, т. е. их реализация облегчалась. По мнению Мэгуна, тормозные влияния на рефлексы спинного мозга может оказывать только РФ продолговатого мозга, а облегчающие влияния регулируются всей РФ ствола и спинного мозга.
Промежуточный мозг
Промежуточный мозг (diencephalon) интегрирует сенсорные, двигательные и вегетативные реакции, необходимые для целостной деятельности организма. Основными образованиями промежуточного мозга являются таламус, гипоталамус, который состоит из свода и эпифиза, и таламической области, которая включает в себя таламус, эпиталамус и метаталамус.
Таламус
Таламус (thalamus, зрительный бугор) — структура, в которой происходит обработка и интеграция практически всех сигналов, идущих в кору большого мозга от спинного, среднего мозга, мозжечка, базальных ганглиев головного мозга.
Морфофункциональная организация. В ядрах таламуса происходит переключение информации, поступающей от экстеро-, проприорецепторов и интероцепторов и начинаются таламокортикальные пути.
Учитывая, что коленчатые тела таламуса являются подкорковыми центрами зрения и слуха, а узел уздечки и переднее зрительное ядро участвуют в анализе обонятельных сигналов, можно утверждать, что зрительный бугор в целом является подкорковой «станцией» для всех видов чувствительности. Здесь раздражения внешней и внутренней среды интегрируются, после чего поступают в кору большого мозга.
Зрительный бугор является центром организации и реализации инстинктов, влечений, эмоций. Возможность получать информацию о состоянии множества систем организма позволяет таламусу участвовать в регуляции и определении функционального состояния организма в целом (подтверждением тому служит наличие в таламусе около 120 разнофункциональных ядер). Ядра образуют своеобразные комплексы, которые можно разделить по признаку проекции в кору на 3 группы: передняя проецирует аксоны своих нейронов в поясную извилину коры большого мозга; медиальная — в лобную долю коры; латеральная — в теменную, височную, затылочную доли коры. По проекциям определяется и функция ядер. Такое деление не абсолютно, так как одна часть волокон от ядер таламуса идет в строго ограниченные корковые образования, другая — в разные области коры большого мозга.
Ядра таламуса функционально по характеру входящих и выходящих из них путей делятся на специфические, неспецифические и ассоциативные.
К специфическим ядрам относятся переднее вентральное, медиальное, вентролатеральиое, постлатеральное, постмедиальное, латеральное и медиальное коленчатые тела. Последние относятся к подкорковым центрам зрения и слуха соответственно.
Основной функциональной единицей специфических таламических ядер являются «релейные» нейроны, у которых мало дендритов и длинный аксон; их функция заключается в переключении информации, идущей в кору большого мозга от кожных, мышечных и других рецепторов.
От специфических ядер информация о характере сенсорных стимулов поступает в строго определенные участки III—IV слоев коры большого мозга (соматотопическая локализация). Нарушение функции специфических ядер приводит к выпадению конкретных видов чувствительности, так как ядра таламуса, как и кора большого мозга, имеют соматотопическую локализацию. Отдельные нейроны специфических ядер таламуса возбуждаются рецепторами только своего типа. К специфическим ядрам таламуса идут сигналы от рецепторов кожи, глаз, уха, мышечной системы. Сюда же конвергируют сигналы от интерорецепторов зон проекции блуждающего и чревного нервов, гипоталамуса.
Латеральное коленчатое тело имеет прямые эфферентные связи с затылочной долей коры большого мозга и афферентные связи с сетчаткой глаза и с передними буграми четверохолмий. Нейроны латеральных коленчатых тел по-разному реагируют на цветовые раздражения, включение, выключение света, т. е. могут выполнять детекторную функцию.
В медиальное коленчатое тело (МТК) поступают афферентные импульсы из латеральной петли и от нижних бугров четверохолмий. Эфферетные пути от медиальных коленчатых тел идут в височную зону коры большого мозга, достигая там первичной слуховой области коры. МКТ имеет четкую тонотопичность. Следовательно, уже на уровне таламуса обеспечивается пространственное распределение чувствительности всех сенсорных систем организма, в том числе сенсорных посылок от интерорецепторов сосудов, органов брюшной, грудной полостей.
Ассоциативные ядра таламуса представлены передним медиодорсальным, латеральным дорсальным ядрами и подушкой. Переднее ядро связано с лимбической корой (поясной извилиной), медиодорсальное — с лобной долей коры, латеральное дорсальное — с теменной, подушка — с ассоциативными зонами теменной и височной долями коры большого мозга.
Основными клеточными структурами этих ядер являются мультиполярные, биполярные трехотростчатые нейроны, т. е. нейроны, способные выполнять полисенсорные функции. Ряд нейронов изменяет активность только при одновременном комплексном раздражении. На полисенсорных нейронах происходит конвергенция возбуждений разных модальностей, формируется интегрированный сигнал, который затем передается в ассоциативную кору мозга. Нейроны подушки связаны главным образом с ассоциативными зонами теменной и височной долей коры большого мозга, нейроны латерального ядра — с теменной, нейроны медиального ядра — с лобной долей коры большого мозга.
Неспецифические ядра таламуса представлены срединным центром, парацентральным ядром, центральным медиальным и латеральным, субмедиальным, вентральным передним, парафасцикулярным комплексами, ретикулярным ядром, перивентрикулярной и центральной серой массой. Нейроны этих ядер образуют свои связи по ретикулярному типу. Их аксоны поднимаются в кору большого мозга и контактируют со всеми ее слоями, образуя не локальные, а диффузные связи. К неспецифическим ядрам поступают связи из РФ ствола мозга, гипоталамуса, лимбической системы, базальных ганглиев, специфических ядер таламуса.
Возбуждение неспецифических ядер вызывает генерацию в коре специфической веретенообразной электрической активности, свидетельствующей о развитии сонного состояния. Нарушение функции неспецифических ядер затрудняет появление веретенообразной активности, т. е. развитие сонного состояния.
Сложное строение таламуса, наличие в нем взаимосвязанных специфических, неспецифических и ассоциативных ядер, позволяет ему организовывать такие двигательные реакции, как сосание, жевание, глотание, смех. Двигательные реакции интегрируются в таламусе с вегетативными процессами, обеспечивающими эти движения.
Конвергенцию сенсорных стимулов в таламус обусловливает возникновение так называемых таламических неукротимых болей, которые возникают при патологических процессах в самом таламусе.
Глава 4. НЕРВНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Мозжечок
Мозжечок (cerebellum, малый мозг) — одна из интегративных структур головного мозга, принимающая участие в координации и регуляции произвольных, непроизвольных движений, в регуляции вегетативных и поведенческих функций.
Особенности морфофункциональной организации и связи мозжечка. Реализация указанных функций обеспечивается следующими морфологическими особенностями мозжечка:
1) кора мозжечка построена достаточно однотипно, имеет стереотипные связи, что создает условия для быстрой обработки информации;
основной нейронный элемент коры — клетка Пуркинье, имеет большое количество входов и формирует единственный аксонный выход из мозжечка, коллатерали которого заканчиваются на ядерных его структурах;
на клетки Пуркинье проецируются практически все видысенсорных раздражений: проприоцептивные, кожные, зрительные,слуховые, вестибулярные и др.;
выходы из мозжечка обеспечивают его связи с корой большого мозга, со стволовыми образованиями и спинным мозгом.
Мозжечок анатомически и функционально делится на старую, древнюю и новую части.
К старой части мозжечка (archicerebellum) — вестибулярный мозжечок — относится клочково-флоккулярная доля. Эта часть имеет наиболее выраженные связи с вестибулярным анализатором, что объясняет значение мозжечка в регуляции равновесия.
Древняя часть мозжечка (paleocerebellum) — спинальный мозжечок — состоит из участков червя и пирамиды мозжечка, язычка, околоклочкового отдела и получает информацию преимущественно от проприорецептивных систем мышц, сухожилий, надкостницы, оболочек суставов.
Новый мозжечок (neocerebellum) включает в себя кору полушарий мозжечка и участки червя; он получает информацию от коры, преимущественно по лобно-мостомозжечковому пути, от зрительных и слуховых рецептирующих систем, что свидетельствует об его участии в анализе зрительных, слуховых сигналов и организации на них реакции.
Кора мозжечка имеет специфическое, нигде в ЦНС не повторяющееся, строение. Верхний (I) слой коры мозжечка — молекулярный слой, состоит из параллельных волокон, разветвлений дендритов и аксонов II и III слоев. В нижней части молекулярного слоя встречаются корзинчатые и звездчатые клетки, которые обеспечивают взаимодействие клеток Пуркинье.
Средний (II) слой коры образован клетками Пуркинье, выстроенными в один ряд и имеющими самую мощную в ЦНС дендритную систему. На дендритном поле одной клетки Пуркинье может быть до 60 000 синапсов. Следовательно, эти клетки выполняют задачу сбора, обработки и передачи информации. Аксоны клеток Пуркинье являются единственным путем, с помощью которого кора мозжечка передает информацию в его ядра и ядра структуры большого мозга.
Под II слоем коры (под клетками Пуркинье) лежит гранулярный (III) слой, состоящий из клеток-зерен, число которых достигает 10 млрд. Аксоны этих клеток поднимаются вверх, Т-образно делятся на поверхности коры, образуя дорожки контактов с клетками Пуркинье. Здесь же лежат клетки Гольджи.
Из мозжечка информация уходит через верхние и нижние ножки. Через верхние ножки сигналы идут в таламус, в мост, красное ядро, ядра ствола мозга, в ретикулярную формацию среднего мозга. Через нижние ножки мозжечка сигналы идут в продолговатый мозг к его вестибулярным ядрам, оливам, ретикулярной формации. Средние ножки мозжечка связывают новый мозжечок с лобной долей мозга.
Импульсная активность нейронов регистрируется в слое клеток Пуркинье и гранулярном слое, причем частота генерации импульсов этих клеток колеблется от 20 до 200 в секунду. Клетки ядер мозжечка генерируют импульсы значительно реже — 1—3 импульса в секунду.
Стимуляция верхнего слоя коры мозжечка приводит к длительному (до 200 мс) торможению активности клеток Пуркинье. Такое же их торможение возникает при световых и звуковых сигналах. Суммарные изменения электрической активности коры мозжечка на раздражение чувствительного нерва любой мышцы выглядят в форме позитивного колебания (торможение активности коры, гиперполяризация клеток Пуркинье), которое наступает через 15— 20 мс и длится 20—30 мс, после чего возникает волна возбуждения, длящаяся до 500 мс (деполяризация клеток Пуркинье).
В кору мозжечка от кожных рецепторов, мышц, суставных оболочек, надкостницы сигналы поступают по так называемым спинно-мозжечковым трактам: по заднему (дорсальному) и переднему (вентральному). Эти пути к мозжечку проходят через нижнюю оливу продолговатого мозга. От клеток олив идут так называемые лазающие волокна, которые ветвятся на дендритах клеток Пуркинье.
Ядра моста посылают афферентные пути в мозжечок, образующие мшистые волокна, которые оканчиваются на клетках-зернах III слоя коры мозжечка. Между мозжечком и голубоватым местом среднего мозга существует афферентная связь с помощью адренергических волокон. Эти волокна способны диффузно выбрасывать норадреналин в межклеточное пространство коры мозжечка, тем самым гуморально изменяют состояние возбудимости его клеток.
Аксоны клеток III слоя коры мозжечка вызывают торможение клеток Пуркинье и клеток-зерен своего же слоя.
Клетки Пуркинье в свою очередь тормозят активность нейронов ядер мозжечка. Ядра мозжечка имеют высокую тоническую активность и регулируют тонус ряда моторных центров промежуточного, среднего, продолговатого, спинного мозга.
Подкорковая система мозжечка состоит из трех функционально разных ядерных образований: ядра шатра, пробковидного, шаровидного и зубчатого ядра.
Ядро шатра получает информацию от медиальной зоны коры мозжечка и связано с ядром Дейтерса и РФ продолговатого и среднего мозга. Отсюда сигналы идут по ретикулоспинальному пути к мотонейронам спинного мозга.
Промежуточная кора мозжечка проецируется на пробковидное и шаровидное ядра. От них связи идут в средний мозг к красному ядру, далее в спинной мозг по руброспинальному пути. Второй путь от промежуточного ядра идет к таламусу и далее в двигательную зону коры большого мозга.
Зубчатое ядро, получая информацию от латеральной зоны коры мозжечка, связано с таламусом, а через него — с моторной зоной коры большого мозга.
Мозжечковый контроль двигательной активности. Эфферентные сигналы из мозжечка к спинному мозгу регулируют силу мышечных сокращений, обеспечивают способность к длительному тоническому сокращению мышц, способность сохранять оптимальный тонус мышц в покое или при движениях, соразмерять произвольные движения с целью этого движения, быстро переходить от сгибания к разгибанию и наоборот.
Мозжечок обеспечивает синергию сокращений разных мышц при сложных движениях. Например, делая шаг при ходьбе, человек заносит вперед ногу, одновременно центр тяжести туловища переносится вперед при участии мышц спины. В тех случаях, когда мозжечок не выполняет своей регуляторной функции, у человека наблюдаются расстройства двигательных функций, что выражается следующими симптомами.
астения (astenia — слабость) — снижение силы мышечного сокращения, быстрая утомляемость мышц;
астазия (astasia, от греч. а — не, stasia — стояние) — утрата способности к длительному сокращению мышц, что затрудняет стояние, сидение и т. д.;
дистония (distonia — нарушение тонуса) — непроизвольное повышение или понижение тонуса мышц;
тремор (tremor — дрожание) — дрожание пальцев рук, кистей, головы в покое; этот тремор усиливается при движении;
дисметрия (dismetria — нарушение меры) — расстройство равномерности движений, выражающееся либо в излишнем, либо недостаточном движении. Больной пытается взять предмет со стола и проносит руку за предмет (гиперметрия) или не доносит ее до предмета (гипометрия);
атаксия (ataksia, от греч. а — отрицание, taksia — порядок) — нарушение координации движений. Здесь ярче всего проявляется невозможность выполнения движений в нужном порядке, в определенной последовательности. Проявлениями атаксии являются так же адиадохокинез, асинергия, пьяная-шаткая походка. При адиадохокинезе человек не способен быстро вращать ладони вниз—вверх. При асинергии мышц он не способен сесть из положения лежа без помощи рук. Пьяная походка характеризуется тем, что человек ходит, широко расставив ноги, шатаясь из стороны в сторону от линии ходьбы. Врожденных двигательных актов у человека не так уж много (например, сосание), большинство же движений он выучивает в течение жизни и они становятся автоматическими (ходьба, письмо и т.д.). Когда нарушается функция мозжечка, движения становятся неточными, негармоничными, разбросанными, часто не достигают цели.
Данные о том, что повреждение мозжечка ведет к расстройствам движений, которые были приобретены человеком в результате обучения, позволяют сделать вывод, что само обучение шло с участием мозжечковых структур, а следовательно, мозжечок принимает участие в организации процессов высшей нервной деятельности;
7) дизартрия (disartria) — расстройство организации речевой моторики. При повреждении мозжечка речь больного становится растянутой, слова иногда произносятся как бы толчками (скандированная речь).
При повреждении мозжечка наблюдается повышение тонуса мышц-разгибателей. Регуляция мышечного тонуса с помощью мозжечка происходит следующим образом: проприоцептивные сигналы о тонусе мышц поступают в область червя и клочково-узелковую долю, отсюда — в ядро шатра, далее — к ядру преддверия и РФ продолговатого и среднего мозга и, наконец, по ретикулярно- и вестибулоспинальным путям к нейронам передних рогов спинного мозга, иннервирующих мышцы, от которых поступили сигналы. Следовательно, регуляция мышечного тонуса реализуется по принципу обратной связи.
Следует отметить, что характер влияния на тонус мышц определяется частотой генерации импульсов нейронов ядра шатра. При высокой частоте (30—300 имп/с) тонус мышц-разгибателей снижается, при низкой (2—10 имп/с) — увеличивается.
Промежуточная область коры мозжечка получает информацию по спинальным трактам от двигательной области коры большого мозга (прецентральной извилины), по коллатералям пирамидного пути, идущего в спинной мозг. Коллатерали заходят в мост, а оттуда — в кору мозжечка. Следовательно, за счет коллатералей мозжечок получает информацию о готовящемся произвольном движении, и возможность участвовать в обеспечении тонуса мышц, необходимого для реализации этого движения.
Латеральная кора мозжечка получает информацию из двигательной области коры большого мозга. В свою очередь латеральная кора посылает информацию в зубчатое ядро мозжечка, отсюда по мозжечково-кортикальному пути в сенсомоторную область коры большого мозга (постцентральная извилина), а через мозжечково-рубральный путь к красному ядру и от него по руброспинальному пути к передним рогам спинного мозга. Параллельно сигналы по пирамидному пути идут к тем же передним рогам спинного мозга.
Таким образом, мозжечок, получив информацию о готовящемся движении, корректирует программу подготовки этого движения в коре и одновременно готовит тонус мускулатуры для реализации этого движения через спинной мозг.
Изменение тонуса мышц после повреждения мозжечка обусловлено тем, что исчезает торможение лабиринтных и миотатических рефлексов, которое в норме осуществляется мозжечком. В норме вестибулярные ядра активируют мотонейроны спинного мозга мышц-разгибателей, а мозжечок тормозит активность нейронов преддверного ядра. При повреждении мозжечка вестибулярные ядра бесконтрольно активируют мотонейроны передних рогов спинного мозга, в результате повышается тонус мышц-разгибателей конечностей.
При повреждении мозжечка усиливаются и проприоцептивные рефлексы спинного мозга (рефлексы, вызываемые при раздражении рецепторов сухожилий, мышц, надкостницы, оболочек суставов), но в этом случае снимается тормозное влияние на мотонейроны спинного мозга ретикулярной формации продолговатого мозга.
В норме мозжечок активирует пирамидные нейроны коры большого мозга, которые тормозят активность мотонейронов спинного мозга. Чем больше мозжечок активирует пирамидные нейроны коры, тем более выражено торможение мотонейронов спинного мозга. При повреждении мозжечка это торможение исчезает, так как активация пирамидных клеток прекращается.
Таким образом, в случае повреждения мозжечка активируются нейроны вестибулярных ядер и ретикулярной формации продолговатого мозга, которые активируют мотонейроны спинного мозга. Одновременно активность пирамидных нейронов снижается, а следовательно, снижается их тормозное влияние на те же мотонейроны спинного мозга. В итоге, получая возбуждающие сигналы от продолговатого мозга при одновременном уменьшении тормозных влияний от коры большого мозга (после повреждения структур мозжечка), мотонейроны спинного мозга активируются и вызывают гипертонус мышц.
Взаимодействие мозжечка и коры большого мозга. Это взаимодействие организовано соматотопически. Функционально мозжечок может оказывать облегчающее, тормозящее и компенсаторное влияние на реализацию функций коры большого мозга.
Роль взаимодействия лобной доли коры большого мозга с мозжечком хорошо проявляется при частичных повреждениях мозжечка. Одномоментное удаление мозжечка приводит к гибели человека, в то же время, если удаляется часть мозжечка, это вмешательство, как правило, не смертельно. После операции частичного удаления мозжечка возникают симптомы его повреждения (тремор, атаксия, астения и т. д.), которые затем исчезают. Если на фоне исчезновения мозжечковых симптомов нарушается функция лобных долей мозга, то мозжечковые симптомы возникают вновь. Следовательно, кора лобных долей большого мозга компенсирует расстройства, вызываемые повреждением мозжечка. Механизм данной компенсации реализуется через лобно-мостомозжечковый тракт.
Мозжечок за счет своего влияния на сенсомоторную область коры может изменять уровень тактильной, температурной, зрительной чувствительности. Оказалось, что повреждение мозжечка снижает уровень восприятия критической частоты мельканий света (наименьшая частота мельканий, при которой световые стимулы воспринимаются не как отдельные вспышки, а как непрерывный свет).
Удаление мозжечка приводит к ослаблению силы процессов возбуждения и торможения, нарушению баланса между ними, развитию инертности. Выработка двигательных условных рефлексов после удаления мозжечка затрудняется, особенно в случаях формирования локальной, изолированной двигательной реакции. Точно так же замедляется выработка пищевых условных рефлексов, увеличивается скрытый (латентный) период их вызова.
Влияние мозжечка на вегетативные функции. Мозжечок оказывает угнетающее и стимулирующее влияние на работу сердечнососудистой, дыхательной, пищеварительной и других систем организма. В результате двойственного влияния мозжечок стабилизирует, оптимизирует функции систем организма.
Сердечно-сосудистая система реагирует на раздражение мозжечка либо усилением (например, прессорные рефлексы), либо снижением этой реакции. Направленность реакции зависит от фона, на котором она вызывается. При раздражении мозжечка высокое кровяное давление снижается, а исходное низкое — повышается. Раздражение мозжечка на фоне учащенного дыхания (гиперпноэ) снижает частоту дыхания. При этом одностороннее раздражение мозжечка вызывает на своей стороне снижение, а на противоположной — повышение тонуса дыхательных мышц.
Удаление или повреждение мозжечка приводит к уменьшению тонуса мускулатуры кишечника, из-за низкого тонуса нарушается эвакуация содержимого желудка и кишечника. Нарушается также нормальная динамика секреции и всасывания в желудке и кишечнике.
Обменные процессы при повреждении мозжечка идут более интенсивно, гипергликемическая реакция (увеличение количества глюкозы в крови) на введение глюкозы в кровь или на прием ее с пищей возрастает и сохраняется дольше, чем в норме, ухудшается аппетит, наблюдается исхудание, замедляется заживление ран, волокна скелетных мышц подвергаются жировому перерождению.
При повреждении мозжечка нарушается генеративная функция, что проявляется в нарушении последовательности процессов родовой деятельности. При возбуждении или повреждении мозжечка мышечные сокращения, сосудистый тонус, обмен веществ и т. д. реагируют так же, как при активации или повреждении симпатического отдела вегетативной нервной системы.
Таким образом, мозжечок принимает участие в различных видах деятельности организма: моторной, соматической, вегетативной, сенсорной, интегративной и т. д. Однако эти функции мозжечок реализует через другие структуры центральной нервной системы. Мозжечок выполняет функцию оптимизации отношений между различными отделами нервной системы, что реализуется, с одной стороны, активацией отдельных центров, с другой — удержанием этой активности в определенных рамках возбуждения, лабильности и т. д. После частичного повреждения мозжечка могут сохраняться все функции организма, но сами функции, порядок их реализации, количественное соответствие потребностям трофики организма нарушаются.
Глава 4. НЕРВНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Лимбическая система...
* * *
Глава 4. НЕРВНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Гиппокамп
Гиппокамп (hippocampus) расположен в глубине височных долей мозга и является основной структурой лимбической системы. Морфологически гиппокамп представлен стереотипно повторяющимися модулями, связанными между собой и с другими структурами.
Модульное строение обусловливает способность гиппокампа генерировать высокоамплитудную ритмическую активность. Связь модулей создает условие циркулирования активности в гиппокампе при обучении. При этом возрастает амплитуда синаптических потенциалов, увеличиваются нейросекреция клеток гиппокампа, число шипиков на дендритах его нейронов, что свидетельствует о переходе потенциальных синапсов в активные. Многочисленные связи гиппокампа со структурами как лимбической системы, так и других отделов мозга определяют его многофункциональность.
Выраженными и специфическими являются электрические процессы в гиппокампе. Активность здесь чаще всего характеризуется быстрыми бета-ритмами (14—30 в секунду) и медленными тета-ритмами (4—7 в секунду).
Если с помощью фармакологических методов в новой коре ослабить десинхронизацию на новое раздражение, то в гиппокампе затрудняется возникновение тета-ритма. Раздражение ретикулярной формации ствола мозга усиливает выраженность тета-ритма в гиппокампе и высокочастотных ритмов в новой коре.
Значение тета-ритма заключается в том, что он отражает реакцию гиппокампа, а тем самым — его участие в ориентировочном рефлексе, реакциях настороженности, повышения внимания, в динамике обучения. Тета-ритм в гиппокампе наблюдается при высоком уровне эмоционального напряжения — страхе, агрессии, голоде, жажде. Вызванная активность в гиппокампе возникает на раздражение различных рецепторов и любой из структур лимбической системы. Разносенсорные проекционные зоны в гиппокампе перекрываются. Это обусловлено тем, что большинство нейронов гиппокампа характеризуется полисенсорностью, т. е. способностью реагировать на световые, звуковые и другие виды раздражений.
Нейроны гиппокампа отличаются выраженной фоновой активностью. В ответ на сенсорное раздражение реагирует до 60% нейронов гиппокампа. Особенность строения гиппокампа, взаимосвязанные модули обусловливают цикл генерирования возбуждения в нем, что выражается в длительной реакции (до 12 с) нейронов на однократный короткий стимул.
Повреждение гиппокампа у человека нарушает память на события, близкие к моменту повреждения (ретроантероградная амнезия). Нарушаются запоминание, обработка новой информации, различие пространственных сигналов. Повреждение гиппокампа ведет к снижению эмоциональности, инициативности, замедлению скорости основных нервных процессов, повышаются пороги вызова эмоциональных реакций.
Глава 4. НЕРВНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Миндалевидное тело
Миндалевидное тело (corpus amygdoloideum), миндалина — подкорковая структура лимбической системы, расположенная в глубине височной доли мозга. Нейроны миндалины разнообразны по форме, функциям и нейрохимическим процессам в них. Функции миндалины связаны с обеспечением оборонительного поведения, вегетативными, двигательными, эмоциональными реакциями, мотивацией условнорефлекторного поведения.
Электрическая активность миндалин характеризуется разноамплитудными и разночастотными колебаниями. Фоновые ритмы могут коррелировать с ритмом дыхания, сердечных сокращений.
Миндалины реагируют многими своими ядрами на зрительные, слуховые, интероцептивные, обонятельные, кожные раздражения, причем все эти раздражения вызывают изменение активности любого из ядер миндалины, т. е. ядра миндалины полисенсорны. Реакция ядра на внешние раздражения длится, как правило, до 85 мс, т. е. значительно меньше, чем реакция на подобные же раздражения новой коры.
Нейроны имеют хорошо выраженную спонтанную активность, которая может быть усилена или заторможена сенсорными раздражениями. Многие нейроны полимодальны и полисенсорны и активируются синхронно с тета-ритмом.
Раздражение ядер миндалевидного тела создает выраженный парасимпатический эффект на деятельность сердечно-сосудистой, дыхательной систем, приводит к понижению (редко к повышению) кровяного давления, урежению сердечного ритма, нарушению проведения возбуждения по проводящей системе сердца, возникновению аритмий и экстрасистолий. При этом сосудистый тонус может не изменяться.
Урежение ритма сокращений сердца при воздействии на миндалины отличается длительным скрытым периодом и имеет длительное последействие.
Раздражение ядер миндалины вызывает угнетение дыхания, иногда кашлевую реакцию.
При искусственной активации миндалины появляются реакции принюхивания, облизывания, жевания, глотания, саливации, изменения перистальтики тонкой кишки, причем эффекты наступают с большим латентным периодом (до 30—45 с после раздражения). Стимуляция миндалин на фоне активных сокращений желудка или кишечника тормозит эти сокращения.
Разнообразные эффекты раздражения миндалин обусловлены их связью с гипоталамусом, который регулирует работу внутренних органов.
Повреждение миндалины у животных снижает адекватную подготовку автономной нервной системы к организации и реализации поведенческих реакций, приводит к гиперсексуальности, исчезновению страха, успокоению, неспособности к ярости и агрессии. Животные становятся доверчивыми. Например, обезьяны с поврежденной миндалиной спокойно подходят к гадюке, вызывавшей ранее у них ужас, бегство. Видимо, в случае повреждения миндалины исчезают некоторые врожденные безусловные рефлексы, реализующие память об опасности.
Глава 4. НЕРВНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Гипоталамус
Гипоталамус (hypothalamus, подбугорье) — структура промежуточного мозга, входящая в лимбическую систему, организующая эмоциональные, поведенческие, гомеостатические реакции организма.
Морфофункциональная организация. Гипоталамус имеет большое число нервных связей с корой большого мозга, подкорковыми узлами, зрительным бугром, средним мозгом, мостом, продолговатым и спинным мозгом.
В состав гипоталамуса входят серый бугор, воронка с нейрогипофизом и сосцевидные тела. Морфологически в нейронных структурах гипоталамуса можно выделить около 50 пар ядер, имеющих свою специфическую функцию. Топографически эти ядра можно объединить в 5 групп: 1) преоптическая группа имеет выраженные связи с конечным мозгом и делится на медиальное и латеральное предоптические ядра; 2) передняя группа, в состав которой входят супраоптическое, паравентрикулярные ядра; 3) средняя группа состоит из нижнемедиального и верхнемедиального ядер; 4) наружная группа включает в себя латеральное гипоталамическое поле и серобугорные ядра; 5) задняя группа сформирована из медиальных и латеральных ядер сосцевидных тел и заднего гипоталамического ядра.
Ядра гипоталамуса имеют мощное кровоснабжение, подтверждением чему служит тот факт, что ряд ядер гипоталамуса обладает изолированным дублирующим кровоснабжением из сосудов артериального круга большого мозга (виллизиев круг). На 1 мм2 площади гипоталамуса приходится до 2600 капилляров, в то время как на той же площади V слоя предцентральной извилины (моторной коры) их 440, в гиппокампе — 350, в бледном шаре — 550, в затылочной доле коры большого мозга (зрительной коре) — 900. Капилляры гипоталамуса высокопроницаемы для крупномолекулярных белковых соединений, к которым относятся нуклеопротеиды, что объясняет высокую чувствительность гипоталамуса к нейровирусным инфекциям, интоксикациям, гуморальным сдвигам.
У человека гипоталамус окончательно созревает к возрасту 13— 14 лет, когда заканчивается формирование гипоталамо-гипофизарных нейросекреторных связей. За счет мощных афферентных связей с обонятельным мозгом, базальными ганглиями, таламусом, гиппокампом, корой большого мозга гипоталамус получает информацию о состоянии практически всех структур мозга. В то же время гипоталамус посылает информацию к таламусу, ретикулярной формации, вегетативным центрам ствола мозга и спинного мозга.
Нейроны гипоталамуса имеют особенности, которые и определяют специфику функций самого гипоталамуса. К этим особенностям относятся чувствительность нейронов к составу омывающей их крови, отсутствие гематоэнцефалического барьера между нейронами и кровью, способность нейронов к нейросекреции пептидов, нейромедиаторов и др.
Роль гипоталамуса в регуляции вегетативных функций. Влияние на симпатическую и парасимпатическую регуляцию позволяет гипоталамусу воздействовать на вегетативные функции организма гуморальным и нервным путями.
Раздражение ядер передней группы сопровождается парасимпатическими эффектами. Раздражение ядер задней группы вызывает симпатические эффекты в работе органов. Стимуляция ядер средней группы приводит к снижению влияний симпатического отдела автономной нервной системы. Указанное распределение функций гипоталамуса не абсолютно. Все структуры гипоталамуса способны в разной степени вызывать симпатические и парасимпатические эффекты. Следовательно, между структурами гипоталамуса существуют функциональные взаимодополняющие, взаимокомпенсирующие отношения.
В целом за счет большого количества связей, полифункциональности структур гипоталамус выполняет интегрирующую функцию вегетативной, соматической и эндокринной регуляции, что проявляется и в организации его ядрами ряда конкретных функций. Так, в гипоталамусе располагаются центры гомеостаза, теплорегуляции, голода и насыщения, жажды и ее удовлетворения, полового поведения, страха, ярости, регуляции цикла бодрствование—сон. Все эти центры реализуют свои функции путем активации или торможеиия автономного (вегетативного) отдела нервной системы, эндокринной системы, структур ствола и переднего мозга. Нейроны ядер передней группы гипоталамуса продуцируют вазопрессин, или антидиуретический гормон (АДГ), окситоцин и другие пептиды, которые по аксонам попадают в заднюю долю гипофиза — нейрогипофиз.
Нейроны ядер срединной группы гипоталамуса продуцируют так называемые рилизинг-факторы (либерины) и ингибирующие факторы (статины), которые регулируют активность передней доли гипофиза — аденогипофиз. В нем образуются такие вещества, как соматотропный, тиреотропный и другие гормоны (см. раздел 5.2.2). Наличие такого набора пептидов в структурах гипоталамуса свидетельствует о присущей им нейросекреторной функции.
Они также обладают детектирующей функцией: реагируют на изменения температуры крови, электролитного состава и осмотического давления плазмы, количества и состав гормонов крови.
Олдс (Olds) описал поведение крыс, которым вживляли электроды в ядра гипоталамуса и давали возможность самостоятельно стимулировать эти ядра. Оказалось, что стимуляция некоторых ядер приводила к негативной реакции. Животные после однократной самостимуляции больше не подходили к педали, замыкающей стимулирующий ток. При самостимуляции других ядер животные нажимали на педаль часами, не обращая внимания на пищу, воду и др.
Исследования Дельгадо (Delgado) во время хирургических операций показали, что у человека раздражение аналогичных участков вызывало эйфорию, эротические переживания. В клинике показано также, что патологические процессы в гипоталамусе могут сопровождаться ускорением полового созревания, нарушением менструального цикла, половой функции.
Раздражение передних отделов гипоталамуса может вызывать у животных пассивно-оборонительную реакцию, ярость страх, а раздражение заднего гипоталамуса вызывает активную агрессию.
Раздражение заднего гипоталамуса приводит к экзофтальму, расширению зрачков, повышению кровяного давления, сужению просвета артериальных сосудов, сокращениям желчного, мочевого пузырей. Могут возникать взрывы ярости с описанными симпатическими проявлениями. Уколы в области гипоталамуса вызывают глюкозурию, полиурию. В ряде случаев раздражение вызывало нарушение теплорегуляции: животные становились пойкилотермными, у них не возникало лихорадочное состояние.
Гипоталамус является также центром регуляции цикла бодрствование — сон. При этом задний гипоталамус активизирует бодрствование, стимуляция переднего вызывает сон. Повреждение заднего гипоталамуса может вызвать так называемый летаргический сон.
Особое место в функциях гипоталамуса занимает регуляция деятельности гипофиза (см. раздел 5.2.2).
В гипоталамусе и гипофизе образуются также нейрорегуляторные пептиды — энкефалины, эндорфины, обладающие морфиноподобным действием и способствующие снижению стресса и т. д.
Глава 4. НЕРВНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Базальные ядра
Базальные (подкорковые) ядра (nuclei basales) головного мозга располагаются под белым веществом внутри переднего мозга, преимущественно в лобных долях. К базальным ядрам относят хвостатое ядро (nucleus caudatus), скорлупу (putamen), ограду (claustrum), бледный шар (globus pallidus).
Хвостатое ядро. Скорлупа
Хвостатое ядро (nucleus caudatus) и скорлупа (putamen) являются эволюционно более поздними, чем бледный шар, образованиями и функционально оказывают на него тормозящее влияние.
Хвостатое ядро и скорлупа имеют сходное гистологическое строение. Их нейроны относятся ко II типу клеток Гольджи, т. е. имеют короткие дендриты, тонкий аксон; их размер до 20 мк. Этих нейронов в 20 раз больше, чем нейронов Гольджи I типа, имеющих разветвленную сеть дендритов и размер около 50 мк.
Функции любых образований головного мозга определяются прежде всего их связями, которых у базальных ядер достаточно много. Эти связи имеют четкую направленность и функциональную очерченность.
Хвостатое ядро и скорлупа получают нисходящие связи преимущественно от экстрапирамидной коры через подмозолистый пучок. Другие поля коры большого мозга также посылают большое количество аксонов к хвостатому ядру и скорлупе.
Основная часть аксонов хвостатого ядра и скорлупы идет к бледному шару, отсюда — к таламусу и только от него — к сенсорным полям. Следовательно, между этими образованиями имеется замкнутый круг связей. Хвостатое ядро и скорлупа имеют также функциональные связи со структурами, лежащими вне этого круга: с черной субстанцией, красным ядром, люисовым телом, ядрами преддверия, мозжечком, γ -клетками спинного мозга.
Обилие и характер связей хвостатого ядра и скорлупы свидетельствуют об их участии в интегративных процессах, организации и регуляции движений, регуляции работы вегетативных органов.
Раздражение поля 8 коры большого мозга вызывает возбуждение нейронов хвостатого ядра, а поля 6 — возбуждение нейронов хвостатого ядра и скорлупы. Одиночное раздражение сенсомоторной области коры большого мозга может вызывать возбуждение или торможение активности нейронов хвостатого ядра. Эти реакции возникают через 10—20 мс, что свидетельствует о прямых и опосредованных связях коры большого мозга с хвостатым ядром.
Медиальные ядра таламуса имеют прямые связи с хвостатым ядром, свидетельством чего служит реакция его нейронов, наступающая через 2—4 мс после раздражения таламуса.
Реакцию нейронов хвостатого ядра вызывают раздражения кожи, световые, звуковые стимулы. -
Во взаимодействиях хвостатого ядра и бледного шара превалируют тормозные влияния. Если раздражать хвостатое ядро, то большая часть нейронов бледного шара тормозится, а меньшая возбуждается. В случае повреждения хвостатого ядра у животного появляется двигательная гиперактивность.
Взаимодействие черного вещества и хвостатого ядра основано на прямых и обратных связях между ними. Установлено, что стимуляция хвостатого ядра усиливает активность нейронов черного вещества. Стимуляция черного вещества приводит к увеличению, а разрушение — к уменьшению количества дофамина в хвостатом ядре. Установлено, что дофамин синтезируется в клетках черного вещества, а затем со скоростью 0,8 мм/ч транспортируется к синапсам нейронов хвостатого ядра. В хвостатом ядре в 1 г нервной ткани накапливается до 10 мкг дофамина, что в 6 раз больше, чем в других отделах переднего мозга, бледном шаре, в 19 раз больше, чем в мозжечке. Благодаря дофамину проявляется растормаживающий механизм взаимодействия хвостатого ядра и бледного шара.
При недостатке дофамина в хвостатом ядре (например, при дисфункции черного вещества) бледный шар растормаживается, активизирует спинно-стволовые системы, что приводит к двигательным нарушениям в виде ригидности мышц.
Кортико-стриарные связи топически локализованы. Так, передние области мозга связаны с головкой хвостатого ядра. Патология, возникающая в одной из взаимосвязанных областей кора — хвостатое ядро, функционально компенсируется сохранившейся структурой.
Хвостатое ядро и бледный шар принимают участие в таких интегративных процессах, как условнорефлекторная деятельность, двигательная активность. Это выявляется при стимуляции хвостатого ядра, скорлупы и бледного шара, деструкции и при регистрации электрической активности.
Прямое раздражение некоторых зон хвостатого ядра вызывает поворот головы в сторону, противоположную раздражаемому полушарию, животное начинает двигаться по кругу, т. е. возникает так называемая циркуляторная реакция.
Раздражение других областей хвостатого ядра и скорлупы вызывает прекращение всех видов активности человека или животного: ориентировочной, эмоциональной, двигательной, пищевой. При этом в коре большого мозга наблюдается медленноволновая активность.
У человека стимуляция хвостатого ядра во время нейрохирургической операции нарушает речевой контакт с больным: если больной что-то говорил, то он замолкает, а после прекращения раздражения не помнит, что к нему обращались. В случаях травм головного мозга с раздражением головки хвостатого ядра у больных отмечается ретро-, антеро- или ретроантероградная амнезия.
У таких животных, как обезьяны, раздражения хвостатого ядра на разных этапах реализации условного рефлекса приводят к торможению выполнения данного рефлекса. Например, если у обезьяны через вживленные электроды раздражать хвостатое ядро перед подачей условного сигнала, то обезьяна не реагирует на сигнал, как будто не слышала его; раздражение ядра после того, как обезьяна на сигнал направляется к кормушке или уже начинает брать пищу из кормушки, приводит к остановке животного, после прекращения раздражения обезьяна, не завершив условной реакции, возвращается на место, т. е. она «забывает», что был раздражающий сигнал (ретроградная амнезия).
Раздражение хвостатого ядра может полностью предотвратить восприятие болевых, зрительных, слуховых и других видов стимуляции. Раздражение вентральной области хвостатого ядра снижает, а дорсальной — повышает слюноотделение.
При стимуляции хвостатого ядра удлиняются латентные периоды рефлексов, нарушается переделка условных рефлексов. Выработка условных рефлексов на фоне стимуляции хвостатого ядра становится невозможной. Видимо, это объясняется тем, что стимуляция хвостатого ядра вызывает торможение активности коры большого мозга.
Ряд подкорковых структур также получает тормозное влияние хвостатого ядра. Так, стимуляция хвостатых ядер вызывала веретенообразную активность в зрительном бугре, бледном шаре, субталамическом теле, черном веществе и др.
Таким образом, специфичным для раздражения хвостатого ядра является преимущественно торможение активности коры большого мозга, подкорковых образований, торможение безусловного и условнорефлекторного поведения.
В то же время при раздражении хвостатого ядра могут появляться некоторые виды изолированных движений. Видимо, хвостатое ядро имеет наряду с тормозящими и возбуждающие структуры.
Выключение хвостатого ядра сопровождается развитием гиперкинезов типа непроизвольных мимических реакций, тремора, атетоза, торсионного спазма, хореи (подергивания конечностей; туловища, как при некоординированном танце), двигательной гиперактивности в форме бесцельного перемещения с места на место.
В случае повреждения хвостатого ядра наблюдаются существенные расстройства высшей нервной деятельности, затруднение ориентации в пространстве, нарушение памяти, замедление роста организма. После двустороннего повреждения хвостатого ядра условные рефлексы исчезают на длительный срок, выработка новых рефлексов затрудняется, общее поведение отличается застойностью, инертностью, трудностью переключений. У обезьян после одностороннего повреждения хвостатого ядра условная реакция восстанавливалась через 30—50 дней, латентные периоды рефлексов удлинялись, появлялись межсигнальные реакции. Двустороннее повреждение приводило к полному торможению условных рефлексов. Видимо, двустороннее повреждение истощает симметричные компенсаторные механизмы.
При воздействиях на хвостатое ядро, помимо нарушений высшей нервной деятельности, отмечаются расстройства движения. Многие авторы отмечают, что у разных животных при двустороннем повреждении полосатого тела появляется безудержное стремление двигаться вперед, при одностороннем — возникают манежные движения.
Несмотря на большое функциональное сходство хвостатого ядра и скорлупы, имеется ряд функций, специфичных для последней.
Эволюционно скорлупа появляется раньше хвостатого ядра (ее зачатки есть уже у рыб).
Для скорлупы характерно участие в организации пищевого поведения: пищепоиска, пищенаправленности, пищезахвата и пищевладения; ряд трофических нарушений кожи, внутренних органов (например, гепатолентикулярная дегенерация) возникает при нарушениях функции скорлупы. Раздражения скорлупы приводят к изменениям дыхания, слюноотделения.
Как упоминалось ранее, раздражение хвостатого ядра тормозит условный рефлекс на всех этапах его реализации. В то же время раздражение хвостатого ядра препятствует угашению условного рефлекса, т. е. развитию торможения; животное перестает воспринимать новую обстановку. Учитывая, что стимуляция хвостатого ядра приводит к торможению условного рефлекса, следовало бы ожидать, что разрушение хвостатого ядра вызовет облегчение условнорефлекторной деятельности. Но оказалось, что разрушение хвостатого ядра также приводит к торможению условнорефлекторной деятельности. Видимо, функция хвостатого ядра не является просто тормозной, а заключается в корреляции и интеграции процессов оперативной памяти. Это подтверждается также тем, что на нейронах хвостатого ядра конвергирует информация различных сенсорных систем, так как большая часть этих нейронов полисенсорна.
Бледный шар
Бледный шар (globus pallidus s. pallidum) имеет преимущественно крупные нейроны Гольджи I типа. Связи бледного шара с таламусом, скорлупой, хвостатым ядром, средним мозгом, гипоталамусом, соматосенсорной системой и др. свидетельствуют об его участии в организации простых и сложных форм поведения.
Раздражение бледного шара с помощью вживленных электродов вызывает сокращение мышц конечностей, активацию или торможение γ-мотонейронов спинного мозга. У больных с гиперкинезами раздражение разных отделов бледного шара (в зависимости от места и частоты раздражения) увеличивало или снижало гиперкинез.
Стимуляция бледного шара в отличие от стимуляции хвостатого ядра не вызывает торможения, а провоцирует ориентировочную реакцию, движения конечностей, пищевое поведение (обнюхивание, жевание, глотание и т.д.).
Повреждение бледного шара вызывает у людей гипомимию, маскообразность лица, тремор головы, конечностей (причем этот тремор исчезает в покое, во сне и усиливается при движениях), монотонность речи. При повреждении бледного шара наблюдается миоклония — быстрые подергивания мышц отдельных групп или отдельных мышц рук, спины, лица.
В первые часы после повреждения бледного шара в остром опыте на животных резко снижалась двигательная активность, движения характеризовались дискоординацией, отмечалось наличие незавершенных движений, при сидении — поникшая поза. Начав движение, животное долго не могло остановиться. У человека с дисфункцией бледного шара затруднено начало движений, исчезают вспомогательные и реактивные движения при вставании, нарушаются содружественные движения рук при ходьбе, появляется симптом пропульсии: длительная подготовка к движению, затем быстрое движение и остановка. Такие циклы у больных повторяются многократно.
Ограда
Ограда (claustrum) содержит полиморфные нейроны разных типов. Она образует связи преимущественно с корой большого мозга.
Глубокая локализация и малые размеры ограды представляют определенные трудности для ее физиологического исследования. Это ядро имеет форму узкой полоски серого вещества, расположенного под корой большого мозга в глубине белого вещества.
Стимуляция ограды вызывает ориентировочную реакцию, поворот головы в сторону раздражения, жевательные, глотательные, иногда рвотные движения. Раздражение ограды тормозит условный рефлекс на свет, мало сказывается на условном рефлексе на звук. Стимуляция ограды во время еды тормозит процесс поедания пищи.
Известно, что толщина ограды левого полушария у человека несколько больше, чем правого; при повреждении ограды правого полушария наблюдаются расстройства речи.
Таким образом, базальные ядра головного мозга являются интегративными центрами организации моторики, эмоций, высшей нервной деятельности, причем каждая из этих функций может быть усилена или заторможена активацией отдельных образований ба-зальных ядер.
Глава 4. НЕРВНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Кора большого мозга
Высшим отделом ЦНС является кора большого мозга (кора больших полушарий). Она обеспечивает совершенную организацию поведения животных на основе врожденных и приобретенных в онтогенезе функций.
Морфофункциональная организация
Кора большого мозга имеет следующие морфофункциональные особенности:
— многослойность расположения нейронов;
— модульный принцип организации;
— соматотопическая локализация рецептирующих систем;
— экранность, т. е. распределение внешней рецепции на плоскости нейронального поля коркового конца анализатора;
— зависимость уровня активности от влияния подкорковых структур и ретикулярной формации;
— наличие представительства всех функций нижележащих структур ЦНС;
— цитоархитектоническое распределение на поля;
— наличие в специфических проекционных сенсорных и моторной системах вторичных и третичных полей с ассоциативными функциями;
— наличие специализированных ассоциативных областей;
— динамическая локализация функций, выражающаяся в возможности компенсаций функций утраченных структур;
— перекрытие в коре большого мозга зон соседних периферических рецептивных полей;
— возможность длительного сохранения следов раздражения;
— реципрокная функциональная взаимосвязь возбудительных и тормозных состояний;
— способность к иррадиации возбуждения и торможения;
— наличие специфической электрической активности.
Глубокие борозды делят каждое полушарие большого мозга на лобную, височную, теменную, затылочную доли и островок. Островок расположен в глубине сильвиевой борозды и закрыт сверху частями лобной и теменной долей мозга.
Кора большого мозга делится на древнюю (archicortex), старую (paleocortex) и новую (neocortex). Древняя кора наряду с другими функциями имеет отношение к обонянию и обеспечению взаимодействия систем мозга. Старая кора включает поясную извилину, гиппокамп. У новой коры наибольшее развитие величины, дифференциации функций отмечается у человека. Толщина новой коры колеблется от 1,5 до 4,5 мм и максимальна в передней центральной извилине.
Функции отдельных зон новой коры определяются особенностями ее структурно-функциональной организации, связями с другими структурами мозга, участием в восприятии, хранении и воспроизведении информации при организации и реализации поведения, регуляции функций сенсорных систем, внутренних органов.
Особенности структурно-функциональной организации коры большого мозга обусловлены тем, что в эволюции происходила кортикализация функций, т. е. передача коре большого мозга функций нижележащих структур мозга. Однако эта передача не означает, что кора берет на себя выполнение функций других структур. Ее роль сводится к коррекции возможных нарушений функций взаимодействующих с ней систем, более совершенного, с учетом индивидуального опыта, анализа сигналов и организации оптимальной реакции на эти сигналы, формирование в своих и в других заинтересованных структурах мозга памятных следов о сигнале, его характеристиках, значении и характере реакции на него. В дальнейшем, по мере автоматизации реакция начинает выполняться подкорковыми структурами.
Общая площадь коры большого мозга человека около 2200 см2, число нейронов коры превышает 10 млрд. В составе коры имеются пирамидные, звездчатые, веретенообразные нейроны.
Пирамидные нейроны имеют разную величину, их дендриты несут большое количество шипиков; аксон пирамидного нейрона, как правило, идет через белое вещество в другие зоны коры или в структуры ЦНС.
Звездчатые клетки имеют короткие хорошо ветвящиеся дендриты и короткий аскон, обеспечивающий связи нейронов в пределах самой коры большого мозга.
Веретенообразные нейроны обеспечивают вертикальные или горизонтальные взаимосвязи нейронов разных слоев коры.
Кора большого мозга имеет преимущественно шестислойное строение
Слой I — верхний молекулярный, представлен в основном ветвлениями восходящих дендритов пирамидных нейронов, среди которых расположены редкие горизонтальные клетки и клетки-зерна, сюда же приходят волокна неспецифических ядер таламуса, регулирующие через дендриты этого слоя уровень возбудимости коры большого мозга.
Слой II — наружный зернистый, состоит из звездчатых клеток, определяющих длительность циркулирования возбуждения в коре большого мозга, т. е. имеющих отношение к памяти.
Слой III — наружный пирамидный, формируется из пирамидных клеток малой величины и вместе со II слоем обеспечивают корко-корковые связи различных извилин мозга.
Слой IV — внутренний зернистый, содержит преимущественно звездчатые клетки. Здесь заканчиваются специфические таламокортикальные пути, т. е. пути, начинающиеся от рецепторов анализаторов.
Слой V — внутренний пирамидный, слой крупных пирамид, которые являются выходными нейронами, аксоны их идут в ствол мозга и спинной мозг.
Слой VI — слой полиморфных клеток, большинство нейронов этого слоя образуют кортико-таламические пути.
Клеточный состав коры по разнообразию морфологии, функции, формам связи не имеет себе равных в других отделах ЦНС. Нейронный состав, распределение нейронов по слоям в разных областях коры различны, что позволило выделить в мозге человека 53 цитоархитектонических поля. Разделение коры большого мозга на цитоархитектонические поля более четко формируется по мере совершенствования ее функции в филогенезе.
У высших млекопитающих в отличие от низших от двигательного 4 поля хорошо дифференцируются вторичные поля 6, 8 и 10, функционально обеспечивающие высокую координацию, точность движений; вокруг зрительного поля 17 — вторичные зрительные поля 18 и 19, участвующие в анализе значения зрительного стимула (организация зрительного внимания, управление движением глаза). Первичные слуховое, соматосенсорное, кожное и другие поля также имеют рядом расположенные вторичные и третичные поля, обеспечивающие ассоциацию функций данного анализатора с функциями других анализаторов. Для всех анализаторов характерен соматотопический принцип организации проекции на кору большого мозга периферических рецептирующих систем. Так, в сенсорной области коры второй центральной извилины имеются участки представительства локализации каждой точки кожной поверхности, в двигательной области коры каждая мышца имеет свою топику (свое место), раздражая которую можно получить движение данной мышцы; в слуховой области коры имеется топическая локализация определенных тонов (тонотопическая локализация), повреждение локального участка слуховой области коры приводит к потере слуха на определенный тон.
Точно так же в проекции рецепторов сетчатки глаза на зрительное поле коры 17 имеется топографическое распределение. В случае гибели локальной зоны поля 17 изображение не воспринимается, если оно падает на участок сетчатки, проецирующийся на поврежденную зону коры большого мозга.
Особенностью корковых полей является экранный принцип их функционирования. Этот принцип заключается в том, что рецептор проецирует свой сигнал не на один нейрон коры, а на поле нейронов, которое образуется их коллатералями и связями. В результате сигнал фокусируется не точка в точку, а на множестве разнообразных нейронов, что обеспечивает его полный анализ и возможность передачи в другие заинтересованные структуры. Так одно волокно, приходящее в зрительную область коры, может активировать зону размером 0,1 мм. Это значит, что один аксон распределяет свое действие на более чем 5000 нейронов.
Входные (афферентные) импульсы поступают в кору снизу, поднимаются к звездчатым и пирамидным клеткам III—V слоев коры. От звездчатых клеток IV слоя сигнал идет к пирамидным нейронам III слоя, а отсюда по ассоциативным волокнам — к другим полям, областям коры большого мозга. Звездчатые клетки поля 3 переключают сигналы, идущие в кору, на пирамидные нейроны V слоя, отсюда обработанный сигнал уходит из коры к другим структурам мозга.
В коре входные и выходные элементы вместе со звездчатыми клетками образуют так называемые колонки — функциональные единицы коры, организованные в вертикальном направлении. Доказательством этого служит следующее: если микроэлектрод погружать перпендикулярно в кору, то на своем пути он встречает нейроны, реагирующие на один вид раздражения, если же микроэлектрод вводить горизонтально по коре, то он встречает нейроны, реагирующие на разные виды стимулов.
Диаметр колонки около 500 мкм и определяется она зоной распределения коллатералей восходящего афферентного таламокортикального волокна. Соседние колонки имеют взаимосвязи, организующие участки множества колонок в организации той или иной реакции. Возбуждение одной из колонок приводит к торможению соседних.
Каждая колонка может иметь ряд ансамблей, реализующих какую-либо функцию по вероятностно-статистическому принципу. Этот принцип заключается в том, что при повторном раздражении в реакции участвует не вся группа нейронов, а ее часть. Причем каждый раз часть участвующих нейронов может быть разной по составу, т. е. формируется группа активных нейронов (вероятностный принцип), среднестатистически достаточная для обеспечения нужной функции (статистический принцип).
Как уже упоминалось, разные области коры большого мозга имеют разные поля, определяющиеся по характеру и количеству нейронов, толщине слоев и т. д. Наличие структурно различных полей предполагает и разное их функциональное предназначение (рис. 4.14). Действительно, в коре большого мозга выделяют сенсорные, моторные и ассоциативные области.
Сенсорные области
Корковые концы анализаторов имеют свою топографию и на них проецируются определенные афференты проводящих систем. Корковые концы анализаторов разных сенсорных систем перекрываются. Помимо этого, в каждой сенсорной системе коры имеются полисенсорные нейроны, которые реагируют не только на «свой» адекватный стимул, но и на сигналы других сенсорных систем.
Кожная рецептирующая система, таламокортикальные пути проецируются на заднюю центральную извилину. Здесь имеется строгое соматотопическое деление. На верхние отделы этой извилины проецируются рецептивные поля кожи нижних конечностей, на средние — туловища, на нижние отделы — руки, головы.
На заднюю центральную извилину в основном проецируются болевая и температурная чувствительность. В коре теменной доли (поля 5 и 7), где также оканчиваются проводящие пути чувствительности, осуществляется более сложный анализ: локализация раздражения, дискриминация, стереогноз.
При повреждениях коры более грубо страдают функции дистальных отделов конечностей, особенно рук.
Зрительная система представлена в затылочной доле мозга: поля 17, 18, 19. Центральный зрительный путь заканчивается в поле 17; он информирует о наличии и интенсивности зрительного сигнала. В полях 18 и 19 анализируются цвет, форма, размеры, качества предметов. Поражение поля 19 коры большого мозга приводит к тому, что больной видит, но не узнает предмет (зрительная агнозия, при этом утрачивается также цветовая память).
Слуховая система проецируется в поперечных височных извилинах (извилины Гешля), в глубине задних отделов латеральной (сильвиевой) борозды (поля 41, 42, 52). Именно здесь заканчиваются аксоны задних бугров четверохолмий и латеральных коленчатых тел.
Обонятельная система проецируется в области переднего конца гиппокампальной извилины (поле 34). Кора этой области имеет не шести-, а трехслойное строение. При раздражении этой области отмечаются обонятельные галлюцинации, повреждение ее ведет к аносмии (потеря обоняния).
Вкусовая система проецируется в гиппокампальной извилине по соседству с обонятельной областью коры (поле 43).
Моторные области
Впервые Фритч и Гитциг (1870) показали, что раздражение передней центральной извилины мозга (поле 4) вызывает двигательную реакцию. В то же время признано, что двигательная область является анализаторной.
В передней центральной извилине зоны, раздражение которых вызывает движение, представлены по соматотопическому типу, но вверх ногами: в верхних отделах извилины — нижние конечности, в нижних — верхние.
Спереди от передней центральной извилины лежат премоторные поля 6 и 8. Они организуют не изолированные, а комплексные, координированные, стереотипные движения. Эти поля также обеспечивают регуляцию тонуса гладкой мускулатуры, пластический тонус мышц через подкорковые структуры.
В реализации моторных функций принимают участие также вторая лобная извилина, затылочная, верхнетеменная области.
Двигательная область коры, как никакая другая, имеет большое количество связей с другими анализаторами, чем, видимо, и обусловлено наличие в ней значительного числа полисенсорных нейронов.
Ассоциативные области
Все сенсорные проекционные зоны и моторная область коры занимают менее 20% поверхности коры большого мозга (см. рис. 4.14). Остальная кора составляет ассоциативную область. Каждая ассоциативная область коры связана мощными связями с несколькими проекционными областями. Считают, что в ассоциативных областях происходит ассоциация разносенсорной информации. В результате формируются сложные элементы сознания.
Ассоциативные области мозга у человека наиболее выражены в лобной, теменной и височной долях.
Каждая проекционная область коры окружена ассоциативными областями. Нейроны этих областей чаще полисенсорны, обладают большими способностями к обучению. Так, в ассоциативном зрительном поле 18 число нейронов, «обучающихся» условнорефлекторной реакции на сигнал, составляет более 60% от числа фоновоактивных нейронов. Для сравнения: таких нейронов в проекционном поле 17 всего 10—12%.
Повреждение поля 18 приводит к зрительной агнозии. Больной видит, обходит предметы, но не может их назвать.
Полисенсорность нейронов ассоциативной области коры обеспечивает их участие в интеграции сенсорной информации, взаимодействие сенсорных и моторных областей коры.
В теменной ассоциативной области коры формируются субъективные представления об окружающем пространстве, о нашем теле. Это становится возможным благодаря сопоставлению соматосенсорной, проприоцептивной и зрительной информации.
Лобные ассоциативные поля имеют связи с лимбическим отделом мозга и участвуют в организации программ действия при реализации сложных двигательных поведенческих актов.
Первой и наиболее характерной чертой ассоциативных областей коры является мультисенсорность их нейронов, причем сюда поступает не первичная, а достаточно обработанная информация с выделением биологической значимости сигнала. Это позволяет формировать программу целенаправленного поведенческого акта.
Вторая особенность ассоциативной области коры заключается в способности к пластическим перестройкам в зависимости от значимости поступающей сенсорной информации.
Третья особенность ассоциативной области коры проявляется в длительном хранении следов сенсорных воздействий. Разрушение ассоциативной области коры приводит к грубым нарушениям обучения, памяти. Речевая функция связана как с сенсорной, так и с двигательной системами. Корковый двигательный центр речи расположен в заднем отделе третьей лобной извилины (поле 44) чаще левого полушария и был описан вначале Даксом (1835), а затем Брока (1861).
Слуховой центр речи расположен в первой височной извилине левого полушария (поле 22). Этот центр был описан Вернике (1874). Моторный и слуховой центры речи связаны между собой мощным пучком аксонов.
Речевые функции, связанные с письменной речью, — чтение, письмо — регулируются ангулярной извилиной зрительной области коры левого полушария мозга (поле 39).
При поражении моторного центра речи развивается моторная афазия; в этом случае больной понимает речь, но сам говорить не может. При поражении слухового центра речи больной может говорить, излагать устно свои мысли, но не понимает чужой речи, слух сохранен, но больной не узнает слов. Такое состояние называется сенсорной слуховой афазией. Больной часто много говорит (логорея), но речь его неправильная (аграмматизм), наблюдается замена слогов, слов (парафазии).
Поражение зрительного центра речи приводит к невозможности чтения, письма.
Изолированное нарушение письма — аграфия, возникает также в случае расстройства функции задних отделов второй лобной извилины левого полушария.
В височной области расположено поле 37, которое отвечает за запоминание слов. Больные с поражениями этого поля не помнят названия предметов. Они напоминают забывчивых людей, которым необходимо подсказывать нужные слова. Больной, забыв название предмета, помнит его назначения, свойства, поэтому долго описывает их качества, рассказывает, что делают этим предметом, но назвать его не может. Например, вместо слова «галстук» больной, глядя на галстук, говорит: «это то, что надевают на шею и завязывают специальным узлом, чтобы было красиво, когда идут в гости».
Распределение функций по областям мозга не является абсолютным. Установлено, что практически все области мозга имеют полисенсорные нейроны, т. е. нейроны, реагирующие на различные раздражения. Например, при повреждении поля 17 зрительной области его функцию могут выполнять поля 18 и 19. Кроме того, разные двигательные эффекты раздражения одного и того же двигательного пункта коры наблюдаются в зависимости от текущей моторной деятельности.
Если операцию удаления одной из зон коры провести в раннем детском возрасте, когда распределение функций еще не жестко закреплено, функция утраченной области практически полностью восстанавливается, т. е. в коре имеются проявления механизмов динамической локализации функций, позволяющих компенсировать функционально и анатомически нарушенные структуры.
Важной особенностью коры большого мозга является ее способность длительно сохранять следы возбуждения.
Следовые процессы в спинном мозге после его раздражения сохраняются в течение секунды; в подкорково-стволовых отделах (в форме сложных двигательно-координаторных актов, доминантных установок, эмоциональных состояний) длятся часами; в коре мозга следовые процессы могут сохраняться по принципу обратной связи в течение всей жизни. Это свойство придает коре исключительное значение в механизмах ассоциативной переработки и хранения информации, накопления базы знаний.
Сохранение следов возбуждения в коре проявляется в колебаниях уровня ее возбудимости; эти циклы длятся в двигательной области коры 3—5 мин, в зрительной — 5—8 мин.
Основные процессы, происходящие в коре, реализуются двумя состояниями: возбуждением и торможением. Эти состояния всегда реципрокны. Они возникают, например, в пределах двигательного анализатора, что всегда наблюдается при движениях; они могут возникать и между разными анализаторами. Тормозное влияние одного анализатора на другие обеспечивает сосредоточенность внимания на одном процессе.
Реципрокные отношения активности очень часто наблюдаются в активности соседних нейронов.
Отношение между возбуждением и торможением в коре проявляется в форме так называемого латерального торможения. При латеральном торможении вокруг зоны возбуждения формируется зона заторможенных нейронов (одновременная индукция) и она по протяженности, как правило, в два раза больше зоны возбуждения. Латеральное торможение обеспечивает контрастность восприятия, что в свою очередь позволяет идентифицировать воспринимаемый объект.
Помимо латерального пространственного торможения, в нейронах коры после возбуждения всегда возникает торможение активности и наоборот, после торможения — возбуждение — так называемая последовательная индукция.
В тех случаях когда торможение не в состоянии сдерживать возбудительный процесс в определенной зоне, возникает иррадиация возбуждения по коре. Иррадиация может происходить от нейрона к нейрону, по системам ассоциативных волокон I слоя, при этом она имеет очень малую скорость — 0,5—2,0 м/с. В другом случае иррадиация возбуждения возможна за счет аксонных связей пирамидных клеток III слоя коры между соседними структурами, в том числе между разными анализаторами. Иррадиация возбуждения обеспечивает взаимоотношение состояний систем коры при организации условнорефлекторного и других форм поведения.
Наряду с иррадиацией возбуждения, которое происходит за счет импульсной передачи активности, существует иррадиация состояния торможения по коре. Механизм иррадиации торможения заключается в переводе нейронов в тормозное состояние под влиянием импульсов, приходящих из возбужденных участков коры, например, из симметричных областей полушарий.
Электрические проявления активности коры большого мозга
Оценка функционального состояния коры большого мозга человека является трудной и до настоящего времени нерешенной задачей. Одним из признаков, косвенно свидетельствующем о функциональном состоянии структур головного мозга, является регистрация в них колебаний электрических потенциалов.
Каждый нейрон имеет заряд мембраны, который при активации уменьшается, а при торможении — чаще увеличивается, т. е. развивается гиперполяризация. Глия мозга также имеет заряд клеток мембран. Динамика заряда мембраны нейронов, глии, процессы, происходящие в синапсах, дендритах, аксонном холмике, в аксоне — все это постоянно изменяющиеся, разнообразные по интенсивности, скорости процессы, интегральные характеристики которых зависят от функционального состояния нервной структуры и суммарно определяют ее электрические показатели. Если эти показатели регистрируются через микроэлектроды, то они отражают активность локального (до 100 мкм в диаметре) участка мозга и называются фокальной активностью.
В случае, если электрод располагается в подкорковой структуре, регистрируемая через него активность называется субкортикограммой, если электрод располагается в коре мозга — кортикограммой. Наконец, если электрод располагается на поверхности кожи головы, то регистрируется суммарная активность как коры, так и подкорковых структур. Это проявление активности называется электроэнцефалограммой (ЭЭГ) (рис. 4.15).
Все виды активности мозга в динамике подвержены усилению и ослаблению и сопровождаются определенными ритмами электрических колебаний. У человека в покое при отсутствии внешних раздражений преобладают медленные ритмы изменения состояния коры мозга, что на ЭЭГ находит отражение в форме так называемого альфа-ритма, частота колебаний которого составляет 8—13 в секунду, а амплитуда — приблизительно 50 мкВ.
Переход человека к активной деятельности приводит к смене альфа-ритма на более быстрый бета-ритм, имеющий частоту колебаний 14—30 в секунду, амплитуда которых составляет 25 мкВ.
Переход от состояния покоя к состоянию сосредоточенного внимания или ко сну сопровождается развитием более медленного тета-ритма (4—8 колебаний в секунду) или дельта-ритма (0,5—3,5 колебаний в секунду). Амплитуда медленных ритмов составляет 100—300 мкВ (см. рис. 4.15).
Когда на фоне покоя или другого состояния мозгу предъявляется новое быстрое нарастающее раздражение, на ЭЭГ регистрируются так называемые вызванные потенциалы (ВП). Они представляют собой синхронную реакцию множества нейронов данной зоны коры.
Латентный период, амплитуда ВП зависят от интенсивности наносимого раздражения. Компоненты ВП, количество и характер его колебаний зависят от адекватности стимула относительно зоны регистрации ВП.
ВП может состоять из первичного ответа или же из первичного и вторичного. Первичные ответы представляют собой двухфазные, позитивно-негативные колебания. Они регистрируются в первичных зонах коры анализатора и только при адекватном для данного анализатора стимуле. Например, зрительная стимуляция для первичной зрительной коры (поле 17) является адекватной (рис. 4.16). Первичные ответы характеризуются коротким латентным периодом (ЛП), двухфазностью колебания: вначале положительная, затем — отрицательная. Первичный ответ формируется за счет кратковременной синхронизации активности близлежащих нейронов.
Вторичные ответы более вариабельны по ЛП, длительности, амплитуде, чем первичные. Как правило, вторичные ответы чаще возникают на сигналы, имеющие определенную смысловую нагрузку, на адекватные для данного анализатора стимулы; они хорошо формируются при обучении.
Межполушарные взаимоотношения
Взаимоотношение полушарий большого мозга определяется как функция, обеспечивающая специализацию полушарий, облегчение выполнения регуляторных процессов, повышение надежности управления деятельностью органов, систем органов и организма в целом.
Роль взаимоотношений полушарий большого мозга наиболее четко проявляется при анализе функциональной межполушарной асимметрии.
Асимметрия в функциях полушарий впервые была обнаружена в XIX в., когда обратили внимание на различные последствия повреждения левой и правой половины мозга.
В 1836 г. Марк Дакс выступил на заседании медицинского общества в Монпелье (Франция) с небольшим докладом о больных, страдающих потерей речи — состояния, известного специалистам под названием афазии. Дакс заметил связь между потерей речи и поврежденной стороной мозга. В его наблюдениях более чем у 40 больных с афазией имелись признаки повреждения левого полушария. Ученому не удалось обнаружить ни одного случая афазии при повреждении только правого полушария. Суммировав эти наблюдения, Дакс сделал следующее заключение: каждая половина мозга контролирует свои, специфические функции; речь контролируется левым полушарием.
Его доклад не имел успеха. Спустя некоторое время после смерти Дакса Брока при посмертном исследовании мозга больных, страдавших потерей речи и односторонним параличом, отчетливо выявил в обоих случаях очаги повреждения, захватившие части левой лобной доли. С тех пор эта зона стала известна как зона Брока; она была им определена, как область в задних отделах нижней лобной извилины.
Проанализировав связь между предпочтением одной из двух рук и речью, он предположил, что речь, большая ловкость в движениях правой руки связаны с превосходством левого полушария у праворуких.
Через 10 лет после публикации наблюдений Брока концепция, известная теперь как концепция доминантности полушарий, стала основной точкой зрения на взаимоотношения двух полушарий мозга.
В 1864 г. английский невролог Джон Джексон писал: «Не так давно редко кто сомневался в том, что оба полушария одинаковы как в физическом, так и в функциональном плане, но теперь, когда благодаря исследованиям Дакса, Брока и других стало ясно, что повреждение одного полушария может вызвать у человека полную потерю речи, прежняя точка зрения стала несостоятельной».
Д. Джексон выдвинул идею о «ведущем» полушарии, которую можно рассматривать как предшественницу концепции доминантности полушарий. «Два полушария не могут просто дублировать друг друга, — писал он, — если повреждение только одного из них может привести к потере речи. Для этих процессов (речи), выше которых ничего нет, наверняка должна быть ведущая сторона». Далее Джексон сделал вывод о том, «что у большинства людей ведущей стороной мозга является левая сторона так называемой воли, и что правая сторона является автоматической».
К 1870 г. и другие исследователи стали понимать, что многие типы расстройств речи могут быть вызваны повреждением левого полушария. К. Вернике нашел, что больные при повреждении задней части височной доли левого полушария часто испытывали затруднения и в понимании речи.
У некоторых больных при повреждении левого, а не правого полушария обнаруживались затруднения при чтении и письме. Считалось также, что левое полушарие управляет и «целенаправленными движениями».
Совокупность этих данных стала основой представления о взаимоотношении двух полушарий. Одно полушарие (у праворуких обычно левое) рассматривалось как ведущее для речи и других высших функций, другое (правое), или «второстепенное», считали находящимся под контролем «доминантного» левого.
Выявленная первой речевая асимметрия полушарий мозга предопределила представление об эквипотенциальности полушарий большого мозга детей до появления речи. Считается, что асимметрия мозга формируется при созревании мозолистого тела.
Концепция доминантности полушарий, согласно которой во всех гностических и интеллектуальных функциях ведущим у «правшей» является левое полушарие, а правое оказывается «глухим и немым», просуществовала почти столетие. Однако постепенно накапливались свидетельства, что представление о правом полушарии как о второстепенном, зависимом, не соответствует действительности. Так, у больных с нарушениями левого полушария мозга хуже выполняются тесты на восприятие форм и оценку пространственных взаимосвязей, чем у здоровых. Неврологически здоровые испытуемые, владеющие двумя языками (английским и идиш), лучше идентифицируют английские слова, предъявленные в правом поле зрения, а слова на идиш — в левом. Был сделан вывод, что такого рода асимметрия связана с навыками чтения: английские слова читаются слева направо, а слова идиш — справа налево.
Почти одновременно с распространением концепции доминантности полушарий стали появляться данные, указывающие на то, что правое, или второстепенное, полушарие также обладает своими особыми способностями. Так, Джексон выступил с утверждением о том, что в задних долях правого мозга локализована способность к формированию зрительных образов.
Повреждение левого полушария приводит, как правило, к низким показателям по тестам на вербальные способности. В то же время больные с повреждением правого полушария обычно плохо выполняли невербальные тесты, включавшие манипуляции с геометрическими фигурами, сборку головоломок, восполнение недостающих частей рисунков или фигур и другие задачи, связанные с оценкой формы, расстояния и пространственных отношений.
Обнаружено, что повреждение правого полушария часто сопровождалось глубокими нарушениями ориентации и сознания. Такие больные плохо ориентируются в пространстве, не в состоянии найти дорогу к дому, в котором прожили много лет. С повреждением правого полушария были связаны также определенные виды агнозий, т. е. нарушений в узнавании или восприятии знакомой информации, восприятии глубины и пространственных взаимоотношений. Одной из самых интересных форм агнозии является агнозия на лица. Больной с такой агнозией не способен узнать знакомого лица, а иногда вообще не может отличать людей друг от друга. Узнавание других ситуаций и объектов, например, может быть при этом не нарушено. Дополнительные сведения, указывающие на специализацию правого полушария, были получены при наблюдении за больными, страдающими тяжелыми нарушениями речи, у которых, однако, часто сохраняется способность к пению. Кроме того, в клинических сообщениях содержались данные о том, что повреждение правой половины мозга может привести к утрате музыкальных способностей, не затронув речевых. Это расстройство, называемое амузией, чаще всего отмечалось у профессиональных музыкантов, перенесших инсульт или другие повреждения мозга.
После того как нейрохирурги осуществили серию операций с комиссуротомией и были выполнены психологические исследования на этих больных, стало ясно, что правое полушарие обладает собственными высшими гностическими функциями.
Существует представление, что межполушарная асимметрия в решающей мере зависит от функционального уровня переработки информации. В этом случае решающее значение придается не характеру стимула, а особенностям гностической задачи, стоящей перед наблюдателем. Принято считать, что правое полушарие специализировано в переработке информации на образном функциональном уровне, левое — на категориальном. Применение такого подхода позволяет снять ряд трудноразрешимых противоречий. Так, преимущество левого полушария, обнаруженное при чтении нотных и пальцевых знаков, объясняется тем, что эти процессы протекают на категориальном уровне переработки информации. Сравнение слов без их лингвистического анализа успешнее осуществляется при их адресации правой гемисфере, поскольку для решения этих задач достаточна переработка информации на образном функциональном уровне.
Межполушарная асимметрия зависит от функционального уровня переработки информации: левое полушарие обладает способностью к переработке информации как на семантическом, так и на перцептивном функциональных уровнях, возможности правого полушария ограничиваются перцептивным уровнем.
В случаях латерального предъявления информации можно выделить три способа межполушарных взаимодействий, проявляющихся в процессах зрительного опознания.
Параллельная деятельность. Каждое полушарие перерабатывает информацию с использованием присущих ему механизмов.
Избирательная деятельность. Информация перерабатывается в «компетентном» полушарии.
Совместная деятельность. Оба полушария участвуют в переработке информации, последовательно играя ведущую роль на тех или иных этапах этого процесса.
Основным фактором, определяющим участие того или иного полушария в процессах узнавания неполных изображений, является то, каких элементов лишено изображение, а именно какова степень значимости отсутствующих в изображении элементов. В случае, если детали изображения удалялись без учета степени их значимости, опознание в большей мере было затруднено у больных с поражениями структур правого полушария. Это дает основание считать правое полушарие ведущим в опознании таких изображений. Если же из изображения удалялся относительно небольшой, но высокозначимый участок, то опознание нарушалось в первую очередь при поражении структур левого полушария, что свидетельствует о преимущественном участии левой гемисферы в опознании подобных изображений.
В правом полушарии осуществляется более полная оценка зрительных стимулов, тогда как в левом оценнваются наиболее существенные, значимые их признаки.
Когда значительное число деталей изображения, подлежащего опознанию, удалено, вероятность того, что наиболее информативные, значимые его участки не подвергнутся искажению или удалению, невелика, а потому левополушарная стратегия опознания значительно ограничена. В таких случаях более адекватной является стратегия, свойственная правому полушарию, основанная на использовании всей содержащейся в изображении информации.
Трудности в реализации левополушарной стратегии в этих условиях усугубляются еще и тем обстоятельством, что левое полушарие обладает недостаточными «способностями» к точной оценке отдельных элементов изображения. Об этом свидетельствуют также исследования, согласно которым оценка длины и ориентации линий, кривизны дуг, величины углов нарушается прежде всего при поражениях правого полушария.
Иная картина отмечается в случаях, когда большая часть изображения удалена, но сохранен наиболее значимый, информативный его участок. В подобных ситуациях более адекватным является способ опознания, основанный на анализе наиболее значимых фрагментов изображения — стратегия, используемая левым полушарием.
В процессе узнавания неполных изображений участвуют структуры как правого, так и левого полушария, причем степень участия каждого из них зависит от особенностей предъявляемых изображений, и в первую очередь от того, содержит ли изображение наиболее значимые информативные элементы. При наличии этих элементов преобладающая роль принадлежит левому полушарию; при их удалении преимущественную роль в процессе опознания играет правое полушарие.
Глава 4. НЕРВНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Координация движений
Термин «координация» происходит от латинского coordinatio — взаимоупорядочение. Под координацией движений понимают процессы согласования активности мышц тела, направленные на успешное выполнение двигательной задачи.
Для центральной нервной системы объектом управления является опорно-двигательный аппарат. Своеобразие скелетно-мышечной системы заключается в том, что она состоит из большого числа звеньев, подвижно соединенных в суставах, допускающих поворот одного звена относительно другого. Суставы могут позволять звеньям поворачиваться относительно одной, двух или трех осей, т. е. обладать одной, двумя или тремя степенями свободы. Чтобы в трехмерном пространстве достичь любой заданной точки (в пределах длины конечности), достаточно иметь двухзвенную конечность с двумя степенями свободы в проксимальном суставе («плече») и одной степенью свободы в дистальном («локтевом»). На самом деле конечности имеют большее число звеньев и степеней свободы. Поэтому, если бы мы захотели решить геометрическую задачу о том, как должны изменяться углы в суставах, для того чтобы рабочая точка конечности переместилась из одного заданного положения в пространстве в другое, мы обнаружили бы, что эта задача имеет бесконечное множество решений. Чтобы кинематическая цепь совершала нужное движение, необходимо исключить те степени свободы, которые для данного движения являются избыточными. Этого можно достичь двумя способами: 1) можно зафиксировать избыточные степени свободы путем одновременной активации антагонистических групп мышц (коактивация); 2) можно связать движения в разных суставах определенными соотношениями, уменьшив, таким образом, число независимых переменных, с которыми должна «иметь дело» центральная нервная система. Такие устойчивые сочетания одновременных движений в нескольких суставах, направленных на достижение единой цели, получили название синергий.
Весьма своеобразны и «двигатели», используемые в живом организме. Скелетные мышцы представляют собой эластомеры с нелинейной зависимостью развиваемой силы от частоты активации. При этом развитие силы автоматически сопровождается изменениями упругости и вязкости мышцы. Кроме того, как известно, напряжение мышцы зависит от ее длины (угла в суставе) и скорости удлинения или укорочения. Сложность управления движениями в суставах при помощи мышц усугубляется еще и тем, что на каждую степень свободы, как правило, приходится больше одной пары мышц. При этом многие мышцы являются двухсуставными, т. е. действуют не на один, а на два сустава. Поэтому, например, сгибание пальцев руки невозможно без одновременной активации разгибателей кисти, препятствующих действию сгибателей пальцев в лучезапястном сочленении.
Формы участия мышц в осуществлении двигательных актов весьма многообразны. Анатомическая классификация мышц (например, сгибатели и разгибатели, синергисты и антагонисты) не всегда соответствуют их функциональной роли в движениях. Так, некоторые двухсуставные мышцы в одном суставе осуществляют сгибание, а в другом — разгибание. Антагонист может возбуждаться одновременно с агонистом для обеспечения точности движения, и его участие помогает выполнять двигательную задачу. В связи с этим, учитывая функциональный аспект координации, в каждом конкретном двигательном акте целесообразно выделить основную мышцу (основной двигатель), вспомогательные мышцы (синергисты), антагонисты и стабилизаторы (мышцы, фиксирующие, не участвующие в движении суставы). Роль мышц не ограничивается генерацией силы, антагонисты и стабилизаторы часто функционируют в режиме растяжения под нагрузкой. Этот режим используется для плавного торможения движений, амортизации толчков.
На конечный результат движения влияют не только силы, развиваемые мышцами, но и силы немышечного происхождения. К ним относятся силы инерции, создаваемые массами звеньев тела, вовлекаемых в движение, а также силы реакции, возникающие в кинематических цепях при смещении любого из звеньев. Движение смещает различные звенья тела друг относительно друга и меняет конфигурацию тела, а следовательно, по ходу движения изменяются моменты упомянутых сил. Вследствие изменения суставных углов меняются и моменты мышечных сил. На ход движения влияет и масса звеньев тела; моменты сил тоже изменяются в процессе движения из-за изменения ориентации звеньев относительно вектора силы тяжести. В практической деятельности человек вступает во взаимодействие с предметами внешнего мира — различными инструментами, перемещаемыми грузами и т. д., и ему приходится преодолевать силы тяжести, упругости, трения, инерции, возникающие в процессе этого взаимодействия. Немышечные силы вмешиваются в процесс движения и делают необходимым непрерывное согласование с ними деятельности мышечного аппарата. Необходимо также нейтрализовывать действие непредвиденных помех движению, которые могут возникать во внешней среде, и оперативно исправлять допущенные в ходе реализации движения ошибки.
В связи с перечисленными особенностями скелетно-мышечной системы и условиями ее взаимодействия с внешним миром, управление движениями оказывается немыслимым без решения задачи согласования активности большого числа мышц.
Характер этого согласования зависит от двигательной задачи. Так, если нужно взять стакан с водой, то для формирования такого движения центральная нервная система должна располагать информацией о положении стакана относительно тела и об исходном положении руки. Однако, поскольку мы хотим, чтобы это движение было успешным, кисть заранее раскрылась на величину, соответствующую размеру стакана, чтобы сгибатели пальцев сжимали стакан с силой, достаточной для предотвращения проскальзывания, чтобы приложенная сила была достаточной для плавного подъема, но не вызывала резкого отрыва, чтобы ориентация стакана в кисти после захвата все время была вертикальной, т. е. чтобы реализация движения соответствовала двигательной задаче, то необходимы не только данные о пространственных соотношениях, но и разные сведения о свойствах объекта манипулирования. Многие из этих сведений не могут быть получены в ходе самого движения посредством обратных связей, а должны быть заложены в программу предстоящего движения на этапе его планирования. Предполагают, что двигательная память содержит обобщенные классы двигательных программ, из числа которых в соответствии с двигательной задачей выбирается нужная. Эта программа модифицируется применительно к конкретной ситуации: однотипные движения могут выполняться быстрее или медленнее, с большей или меньшей амплитудой. Одна и та же программа может быть реализована разными наборами мышц. В качестве примера на рис. 4.17 приведены образцы почерка при написании слова «координация» правой и левой рукой, а также карандашом, зажатым в зубах, или прикрепленным к носку ботинка. Размышление над этим примером приводит нас к важному выводу о том, что уровень планирования движения и уровень его исполнения не совпадают, иначе говоря, система управления движениями является многоуровневой. Действительно, произвольное пространственно ориентированное движение планируется в терминах трехмерного евклидового пространства: вверх—вниз, вперед—назад, вправо—влево. Для выполнения этого плана необходимо перевести планируемые линейные перемещения в соответствующие угловые переменные (изменения суставных углов), определить, какие мышечные моменты необходимы для этих угловых перемещений, и, наконец, сформировать двигательные команды, которые вызовут активацию мышц, дающую необходимые значения моментов.
В планировании, преобразовании и исполнении двигательной программы участвуют различные структуры нервной системы, организованные по иерархическому принципу. Двигательная программа может быть реализована различными способами. В простейшем случае центральная нервная система посылает заранее сформированную последовательность команд к мышцам, не подвергающуюся во время реализации никакой коррекции. В этом случае говорят о разомкнутой системе управления. Такой способ управления используется при осуществлении быстрых, так называемых баллистических движений. Чаще всего ход осуществления движения сравнивается с его планом на основе сигналов, поступающих от многочисленных рецепторов, и в реализуемую программу вносятся необходимые коррекции — это замкнутая система управления с обратными связями. Однако и такое управление имеет свои недостатки. Вследствие относительно малой скорости проведения сигналов, значительных задержек в центральном звене обратной связи и времени, необходимых для развития усилия мышцей после прихода активирующей посылки, коррекция движения по сигналу обратной связи может запаздывать. Поэтому во многих случаях целесообразно реагировать не на отклонение от плана движения, а на само внешнее возмущение еще до того, как оно успело вызвать это отклонение. Такое управление называют управлением по возмущению.
В осуществлении координации движений участвуют все отделы центральной нервной системы — от спинного мозга до коры большого мозга. У человека двигательные функции достигли наивысшей сложности в результате перехода к прямостоянию и прямохождению (что осложнило задачу поддержания равновесия), специализации передних конечностей для совершения тонких движений, использования двигательного аппарата для коммуникации (речь, письмо). В управление движениями человека включены высшие формы деятельности мозга, связанные с сознанием, что дало основание называть соответствующие движения произвольными.
На спинальном уровне протекают лишь простейшие координации, тем не менее спинной мозг может осуществлять довольно обширные функции, вплоть до «спинального шагания» у животных (Ч. Шеррингтон). Нервные механизмы ствола мозга существенно обогащают двигательный репертуар, обеспечивая координацию правильной установки тела в пространстве за счет шейных и лабиринтных рефлексов (Р. Магнус) и нормального распределения мышечного тонуса. Важная роль в координации движений принадлежит мозжечку. Такие качества движения, как плавность, точность, необходимая сила, реализуются с участием мозжечка путем регуляции временных, скоростных и пространственных характеристик движения. Животные с удаленными полушариями, но с сохраненным стволом мозга по координации движений почти не отличаются от интактных. Полушария мозга (кора и базальные ядра) обеспечивают наиболее тонкие координации движений: двигательные реакции, приобретенные в индивидуальной жизни. Осуществление этих реакций базируется на работе рефлекторного аппарата ствола мозга и спинного мозга, функционирование которых многократно обогащается деятельностью высших отделов центральной нервной системы.
Механизмы координации движений, роль тех или иных отделов ЦНС в управлении движениями изучаются чаще в опытах на животных, однако объектом исследования естественных движений является преимущественно человек, что обусловлено двумя обстоятельствами. Во-первых, человек в зависимости от задачи исследования может воспроизводить любую требуемую форму двигательной деятельности. Во-вторых, движения человека являются проявлением его поведения и трудовой деятельности и поэтому представляют особый интерес как с теоретической точки зрения вследствие их сложности и дифференцированности, так и с практической — в связи с их значением для медицины, физиологии труда, космонавтики, эргономики, физиологии спорта.
Методы исследования движений человека. Многообразие методических приемов, используемых при изучении координации движений человека, можно разделить на две группы. Одна группа методов ориентирована на получение сведений о процессах, лежащих в основе координации движений, путем регистрации внешних двигательных проявлений. Другие методы связаны с непосредственной регистрацией управляющих сигналов, поступающих к мышцам в процессе двигательной активности (электромиография), с регистрацией афферентных сигналов (микронейронография), изменений ЭЭГ, предшествующих началу движения.
Для регистрации механических параметров движения — траекторий, скорости, ускорения, развиваемой силы (механограмм) используют технику превращения неэлектрических величин в электрические с помощью различных датчиков. Так, с помощью тензо-датчиков можно непосредственно измерять и регистрировать силу, прилагаемую к тому или иному инструменту, или реакции опоры при ходьбе, с помощью резистивных датчиков на основе потенциометров — регистрировать изменения суставных углов при движении. Дифференцирование сигналов с помощью электронных дифференциаторов или ЭВМ позволяет одновременно с записью перемещения получать запись скорости и ускорения.
При анализе механических параметров движения может быть использован принцип циклографии — регистрации последовательных моментов движения путем фотосъемки движущегося человека через равные промежутки времени на неподвижную пластинку. Если на суставах или точках, соответствующих положению центров тяжести звеньев тела, укреплены светящиеся лампочки, то по такой записи — циклограмме, можно восстановить траекторию движения. В настоящее время чаще используются специализированные компьютерные системы регистрации движения с непосредственным вводом в ЭВМ изображений с двух телекамер, образующих стереопару. Решая обратную задачу механики по записям кинематических параметров (если известно распределение масс), можно рассчитать возникающие при движении силы, моменты в суставах, работу и мощность.
Движения глаз можно регистрировать электрически (электроокулография), с помощью контактных линз (внутри них располагается виток проволоки, в котором наводится напряжение при повороте витка в магнитном поле) и другими методами.
Анализ работы мышц при совершении двигательного акта осуществляется с помощью электромиографии. При электромиографическом исследовании движений обычно используют накожные электроды, укрепляемые над исследуемой мышцей. Многоканальный электромиограф дает возможность одновременно записывать электромиограммы нескольких мышц. Амплитуда электромиографического сигнала возрастает с увеличением развиваемой мышцей силы, поэтому электромиограммы в сопоставлении с регистрируемыми синхронно механограммами позволяют судить о силе сокращения мышц и о распределении их активности в последовательных фазах двигательного акта.
Набор средств изучения нейронных механизмов управления движениями на человеке включает регистрацию сухожильных рефлексов и рефлекса Гофмана (Н-рефлекс), дающих оценку уровня возбудимости двигательных нейронов спинного мозга в разные фазы движения. Позже стала возможной регистрация через кожу головы электрических потенциалов различных областей головного мозга, сопутствующих или предшествующих движению (потенциалы готовности). В последние годы разработаны методы неповреждающей стимуляции отдельных областей коры мозга человека с помощью чрезвычайно кратковременных импульсов высокого напряжения (электрическая стимуляция) или кратковременных и очень сильных магнитных полей, создаваемых индуктивными катушками, укрепленными над разными областями головы и индуцирующими электрические поля, достаточные для стимуляции мозговых структур (магнитная стимуляция).
Физиология движений изучает роль сигналов от различных рецепторов в планировании и осуществлении движений и поддержании позы. Среди них есть как простейшие — закрывание глаз, так и более сложные — использование призматических очков, смещающих или переворачивающих изображения окружающего мира, специальных систем, позволяющих вызвать у человека иллюзию движения зрительного окружения. Значение вестибулярного аппарата можно изучать в условиях его гальванической или калорической стимуляции, а также при искусственном изменении величины и направления вектора силы тяжести — на центрифуге, в условиях кратковременной или длительной невесомости. Поступающие от мышцы проприоцептивные сигналы можно менять, прикладывая к ее сухожилию вибрацию, вызывающую активацию рецепторов мышечных веретен.
С помощью описанных методов изучены такие сложные естественные двигательные акты, как ходьба, бег, рабочие и спортивные движения. Многие из этих методов используются в клинике при исследованиях нарушений двигательной функции вследствие поражения нервной системы или опорно-двигательного аппарата.
Ходьба и бег. Ходьба является наиболее распространенной формой локомоции человека (локомоция — активное перемещение в пространстве на расстояния, значительно превышающие размеры тела). Она относится к циклическим двигательным актам, при которых последовательные фазы движения периодически повторяются.
Для удобства изучения и описания цикл ходьбы подразделяют на фазы: для каждой ноги выделяют фазу опоры, в течение которой нога контактирует с опорой, и фазу переноса, когда нога находится в воздухе. Фазы опоры двух ног частично перекрываются по времени, образуя двухопорный период. Центр масс тела человека при ходьбе совершает сложные пространственные движения. Амплитуда этих движений составляет около 5 см в направлении вверх—вниз и 2—4 см в боковом направлении. Наиболее низкое положение центра масс соответствует двухопорному периоду, а наиболее высокое — середине одноопорного периода. Давление на опору во время ходьбы непостоянно. Оно превышает вес тела во время динамических толчков и меньше веса тела в середине одноопорного периода. Ходьба здорового человека характеризуется симметрией движений левой и правой сторон. Фаза опоры начинается с момента соприкосновения пятки с опорой, нога при этом полностью разогнута. Перенос нагрузки на эту ногу сопровождается подошвенным сгибанием стопы и подгибанием в коленном суставе. За счет этих угловых движений в начале фазы опоры происходит «прислаивание» стопы к опоре. В записи опорных реакций в это время отмечают первый максимум — передний динамический толчок. Задний динамический толчок соответствует отталкиванию задней ноги от опоры и осуществляется главным образом мышцами голеностопного сустава и в меньшей степени коленного и тазобедренного суставов. В осуществлении ходьбы участвуют также мышцы спины (рис. 4.18). Наибольшая активность мышц наблюдается в фазу опоры. При этом за счет одновременной активности сгибателей и разгибателей нога становится опорной, способной нести вес тела.
Сопоставление изменений межзвенных узлов в тазобедренном, коленном и голеностопном суставах с распределением по времени активности мышц приводит к выводу, что движение ноги в фазу переноса в значительной степени осуществляется за счет сил инерции подобно двухзвенному маятнику.
Анализ работы мышц при ходьбе показывает, что в разные фазы шага они сокращаются в разных режимах — концентрическом, т. е. с укорочением (мышцы, осуществляющие подошвенное сгибание стопы в опорном периоде), эксцентрическом, т. е. с удлинением (передняя большеберцовая мышца во время «прислаивания» стопы обеспечивает плавность ее опускания на опору), изометрическом, т. е. без изменения длины (мышцы тазобедренного сустава во время переката через пятку).
Повторяемость параметров движений в последовательных циклах при ходьбе не абсолютная: движения обладают некоторой вариабельностью. Наименьшая вариабельность у кинематической картины ходьбы, наибольшая — в работе мышц, проявляющаяся в изменениях электромиограмм от цикла к циклу. Это отражает корригирующую деятельность ЦНС, которая при каждом шаге вносит в стандартную структуру ходьбы поправки, необходимые для обеспечения относительного постоянства ее кинематики.
Бег отличается от ходьбы тем, что нога, которая находится позади, отталкивается от опоры раньше, чем другая опускается на эту опору. В результате в беге имеется безопорный период — период полета. В беге благодаря большим, чем при ходьбе, скоростям перемещения более значительную роль играют баллистические компоненты движения — перемещение звеньев ноги по инерции.
Рабочие движения. Рабочими движениями в широком смысле слова могут быть названы самые разные целенаправленные движения, совершаемые как в процессе труда, так и в повседневной жизни.
У человека основным рабочим органом является рука, причем для выполнения двигательной задачи обычно наиболее важно положение кисти, которая в результате движения должна в определенный момент оказаться в определенном месте пространства. Благодаря большому числу степеней свободы верхней конечности кисть может попасть в нужную точку по разным траекториям и при различных соотношениях углов в плечевом, локтевом и лучезапястном суставах. Это многообразие возможностей позволяет выполнять двигательную задачу, начиная движение из различных исходных поз, и в то же время ставит ЦНС перед сложной задачей выбора одного варианта из многих.
Электромиографические исследования ряда рабочих движений показали сложную картину работы мышц, однако в этой картине часто можно выделить устойчивые сочетания активности некоторых мышц, используемые в различных движениях. Это синергии, основанные на врожденных или выработанных в процессе опыта связях, которые, являясь устойчивыми компонентами движений, упрощают управление сложными двигательными актами и помогают преодолеть избыточность числа мышц и количества степеней свободы.
При совершении одного и того же, даже простого, движения организация мышечной деятельности в сильной степени зависит от вмешательства немышечных сил, в частности, внешних по отношению к человеку. Так, при ударе молотком, когда к массе предплечья добавляется масса молотка, и, следовательно, возрастает роль инерции, разгибание предплечья совершается по типу баллистического движения — мышцы-разгибатели активны только в начале разгибания, которое дальше совершается по инерции, а в конце притормаживается мышцами-антагонистами. Аналогичное по кинематике движение при работе напильником (рис. 4.19), когда основная внешняя сила — трение, совершается путем непрерывной активности мышцы на протяжении всего разгибания. Если первое из этих двух движений является в основном предпрограммированным, то во втором велика роль обратных связей.
Поза. У млекопитающих животных и человека поддержание позы обеспечивается теми же фазическими мышцами, что и движения, специализированные тонические мышцы отсутствуют. Отличие заключается в том, что при «позной» деятельности сила сокращения мышц обычно невелика, режим близок к изометрическому, длительность сокращения значительна. В «позный», или постуральный, режим работы мышц вовлекаются преимущественно низкопороговые, медленные, устойчивые к утомлению двигательные единицы.
Одна из основных задач «позной» деятельности мышц — удержание нужного положения звеньев тела в поле силы тяжести (удержание головы от свисания, голеностопных суставов от тыльного сгибания при стоянии и др.). Кроме того, «позная» активность может быть направлена на фиксацию суставов, не принимающих участия в осуществляемом движении. В трудовой деятельности удержание позы бывает связано с преодолением внешних сил.
Типичный пример позы — стояние человека. Сохранение равновесия при стоянии возможно в том случае, если проекция центра тяжести тела находится в пределах опорного контура, т. е. площади, занимаемой на плоскости опоры стопами (рис. 4.20). Вертикаль, опущенная из общего центра тяжести тела, при стоянии проходит несколько впереди оси голеностопных и коленных суставов и несколько позади оси тазобедренных. Следовательно, на эти суставы действуют моменты сил тяжести вышерасположенных звеньев тела, а это делает необходимым для удержания позы стояния напряжение многих мышц туловища и ног. Развиваемая этими мышцами сила невелика. Максимальное напряжение при стоянии развивают мышцы голеностопного сустава, меньшее — мышцы коленного и тазобедренного суставов. У большинства мышц активность поддерживается на более или менее постоянном уровне. Другие, например передняя большеберцовая мышца, активируются периодически. Последнее обусловлено небольшими колебаниями центра тяжести тела, постоянно происходящими при стоянии как в сагиттальной, так и во фронтальной плоскостях. Эти колебания могут быть зарегистрированы специальным прибором — стабилографом, работающим с помощью тензодатчиков (рис. 4.21). Мышцы голени противодействуют отклонениям тела, возвращая его в вертикальное положение. Таким образом, поддержание позы — это активный процесс, осуществляющийся, как и движение, с участием обратных связей от рецепторов. В поддержании вертикальной позы участвуют зрение и вестибулярный аппарат. Важную роль играет проприорецепция.
Поза стояния у человека энергетически относительно экономна, так как моменты силы тяжести невелики вследствие близости проекции тяжести тела к осям основных суставов ног. Менее экономно стояние у многих животных, например у кошки и собаки, которые стоят на полусогнутых конечностях.
Поддержание равновесия при стоянии — только частный случай «позной» активности. Механизмы поддержания равновесия используются также при локомоции и повседневной двигательной деятельности. Например, быстрое движение руки может вызывать нарушение равновесия. Обычно этого не происходит, потому что произвольному движению предшествуют такие изменения в системе регуляции позы, которые заранее изменяют распределение «позной» активности мышц и тем самым обеспечивают нейтрализацию последствий движения — так называемые позные компоненты произвольного движения. Эта упреждающая «позная» активность осуществляется автоматически с очень короткими центральными задержками. Роль упреждающей активности в стабилизации положения звеньев тела можно проиллюстрировать простым примером: студент удерживает на ладони вытянутой руки учебник физиологии. Если его товарищ внезапно снимет эту книгу с ладони, то рука резко подпрыгнет вверх, если же студент сам снимет книгу свободной рукой, то ладонь останется на прежнем уровне.
К понятию позы примыкает понятие мышечного тонуса. Термин «тонус» многозначен, в применении к скелетным мышцам им обозначают комплекс явлений. В покое мышечные волокна обладают тургором (упругостью), определяющим их сопротивление давлению и растяжению. Тургор составляет тот компонент тонуса, который не связан со специфической нервной активацией мышцы, обусловливающей ее сокращение. Однако в естественных условиях большинство мышц обычно в некоторой степени активируется нервной системой, в частности, для поддержания позы («позный тонус»). Полное расслабление мышцы, когда электромиографическим методом не регистрируется никаких потенциалов действия, достигается только в условиях полного покоя и при исключении задачи поддержания позы в поле силы тяжести (исследуемая часть тела лежит на опоре).
Другой важный компонент тонуса — рефлекторный, определяется рефлексом на растяжение. При исследовании на человеке он выявляется по сопротивлению растяжения мышцы в случае пассивного поворота звена конечности в суставе. Если в процессе такого исследования записать электромиограмму, то в растягиваемой мышце регистрируется электрическая активность, свидетельствующая об активации двигательных единиц. У здорового человека рефлекс на растяжение при пассивном движении наблюдается только в процессе самого растяжения и притом в случае достаточно большой скорости растяжения. Тонический компонент рефлекса на растяжение, т. е. активность в растянутой мышце, обычно отсутствует, о чем можно судить по отсутствию электромиографического сигнала.
Выработка двигательных навыков. Совершенствование двигательной функции человека в процессе онтогенеза происходит как вследствие продолжающегося в первые годы после рождения созревания отделов нервной системы и врожденных механизмов, участвующих в координации движений, так и в результате обучения, т. е. формирования новых связей, ложащихся в основу программ тех или иных конкретных двигательных актов. Координация новых, непривычных движений имеет характерные черты, отличающие ее от координации тех же движений после обучения.
Обилие степеней свободы в опорно-двигательном аппарате, влияние на результат движения сил тяжести и инерции осложняют выполнение любой двигательной задачи. На первых порах обучения нервная система справляется с этими трудностями, нейтрализуя помехи путем развития дополнительных мышечных напряжений. Мышечный аппарат жестко фиксирует суставы, не участвующие в движении, активно тормозит инерцию быстрых движений. Такой путь преодоления помех, возникающих в ходе движения, энергетически невыгоден и утомителен. Использование обратных связей еще несовершенно — коррекционные посылки, возникающие на их основе, несоразмерны и вызывают необходимость повторных дополнительных коррекций.
На электромиограммах видно, что мышцы-антагонисты даже тех суставов, в которых совершаются движения, активируются одновременно, при этом в циклических движениях мышцы почти не расслабляются. Возбуждены также многие мышцы, не имеющие прямого отношения к данному двигательному акту. Движения, совершаемые в таких условиях, напряжены и неэстетичны (например, движения человека, впервые вышедшего на коньках на лед).
Как показал в своих исследованиях Н. А. Бернштейн, по мере обучения вырабатывается такая структура двигательного акта, при которой немышечные силы включаются в его динамику, становятся составной частью двигательной программы. Излишние мышечные напряжения при этом устраняются, движение становится более устойчивым к внешним возмущениям. На электромиограммах видна концентрация возбуждения мышц во времени и пространстве, периоды активности работающих мышц укорачиваются, а количество мышц, вовлеченных в возбуждение, уменьшается. Это приводит к повышению экономичности мышечной деятельности, а движения делаются более плавными, точными и непринужденными (см. рис. 4.19).
Важную роль в обучении движениям играет рецепция, особенно проприорецепция. В процессе двигательного обучения обратные связи используются не только для коррекции движения по его ходу, но и для коррекции программы следующего движения на основе ошибок предыдущего.
Утомление. При длительной физической работе наступает утомление, которое, в частности, проявляется в изменении координации мышечной деятельности. Возбуждение каждой работающей мышцы становится менее локализованным во времени. В работу вовлекаются другие мышцы, сначала синергисты, компенсирующие снижение силы основных мышц, а затем, по мере нарастания дискоординации — и другие мышцы, в частности антагонисты. Движения становятся менее точными, темп их замедляется.
Картина мышечной активности во время движений, совершаемых на фоне утомления, во многом напоминает картину, наблюдаемую при выполнении новых, непривычных движений.
Нарушения координации движений. Поскольку в управлении движениями принимают участие многие отделы ЦНС, нарушения координации движений могут быть использованы в целях диагностики. Они проявляются нарушениями устойчивости при стоянии и ходьбе, асимметрией движений правой и левой стороны, нарушениями точности движений, снижением силы и уменьшением скорости. Регистрация пространственных и временных характеристик движений с их количественным представлением дает возможность оценить степень двигательных расстройств при различных заболеваниях, ход восстановления двигательных функций, предложить эффективные методы двигательной реабилитации.
Глава 4. НЕРВНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
ФИЗИОЛОГИЯ АВТОНОМНОЙ (ВЕГЕТАТИВНОЙ) НЕРВНОЙ СИСТЕМЫ
Согласно Международной анатомической номенклатуре, термин «автономная нервная система» заменил все ранее существовавшие — «растительная», «висцеральная», «непроизвольная», «вегетативная». Анатомически автономная нервная система представлена ядерными образованиями, лежащими в головном и спинном мозге, нервными ганглиями и нервными сплетениями, иннервирующими гладкую мускулатуру всех органов, сердце и железы. Главная функция автономной нервной системы состоит в поддержании постоянства внутренней среды, или гомеостаза, при различных воздействиях на организм. Наряду с этим автономная нервная система регулирует также деятельность и других органов, которые не участвуют непосредственно в поддержании гомеостаза (внутриглазные мышцы, половые органы). Выделяя регуляцию автономной нервной системой висцеральных функций, следует заметить, что в целостных реакциях организма сенсорные, моторные, соматические и висцеральные компоненты между собой тесно связаны. Специальными исследованиями К. М. Быкова, В. Н. Черниговского и др. показана также возможность условнорефлекторной регуляции висцеральных процессов. Это означает, что высшие отделы головного мозга могут регулировать работу иннервированных автономной нервной системой органов, а также координировать их деятельность в соответствии с текущими потребностями организма.
Функциональная структура автономной нервной системы
На основании структурно-функциональных свойств автономную нервную систему принято делить на симпатическую, парасимпатическую и метасимпатическую части. Из них первые две имеют центральные структуры и периферический нервный аппарат, метасимпатическая же часть целиком лежит на периферии в стенках внутренних органов.
Дуга автономного рефлекса (рис. 4.22), как и соматическая рефлекторная дуга, состоит из трех звеньев: чувствительного (афферентного, сенсорного), ассоциативного (вставочного) и эффекторного. В зависимости от уровня замыкания, т. е. расположения ассоциативного звена, различают местные, или ганглионарные, спинальные, бульварные и т. д. рефлекторные дуги. Рефлексы, возникающие при раздражении чувствительных волокон, идущих в составе симпатических и парасимпатических нервов, вовлекают в деятельность не только автономную, но и соматическую нервную систему. Чувствительные волокна этой единой (автономной и соматической) афферентной системы являются отростками биполярных клеток, лежащих в спинномозговых узлах или их аналогах, таких как яремный, тройничный (гассеров) узлы и др. Такое понимание справедливо для сегментарных и рефлекторных дуг более высокого порядка и не относится к местным периферическим дугам автономного рефлекса.
Наряду с общим для обеих (автономной и соматической) систем звеном существует и собственный афферентный путь автономной нервной системы, называемый особым, или висцеральным. Он создает основу для путей местных рефлексов, осуществляемых независимо, без участия ЦНС. По локализации клеточных тел чувствительных нейронов, по ходу и длине отростков их разделяют на три группы. В первую группу объединены клетки, тела которых локализуются в узлах солнечного и нижнего брыжеечного сплетений. Один из их длинных отростков направляется на периферию, другой в сторону спинного мозга. Клетки второй группы характеризуются тем, что их длинный отросток идет к рабочему органу, короткие распределяются в самом ганглии и синаптически контактируют с вставочным или эффекторным нейронами. Висцеральные чувствительные клетки третьей группы отличаются тем, что их тела и короткие отростки располагаются в интрамуральных узлах, длинные же отростки в составе соответствующих нервов достигают симпатических узлов, где и происходит переключение на ассоциативный и моторный (эфферентный) нейрон.
Висцеральная чувствительность обусловлена активностью пяти отдельных типов интероцепторов: механо-, хемо-, термо-, осмо- и ноцицепторов, называемых специфическими. Из них наиболее распространенными являются механорецепторы.
Среди механорецепторов внутренних органов известны рецепторы двух типов: быстро- и медленноадаптирующиеся. Быстроадаптирующиеся механорецепторы характеризуются высоким порогом возбуждения и встречаются в основном в слизистой оболочке и серозном слое висцеральных органов и связаны преимущественно с миелиновыми волокнами. Характерной чертой быстроадаптирующихся рецепторов являются исключительная чувствительность к динамической фазе движения и сокращения. Для медленноадаптирующихся механорецепторов, наоборот, характерна генерация сигналов в течение длительного периода раздражения или после его окончания. Эти рецепторы имеются во всех внутренних органах и характеризуются низким порогом возбуждения. Такая особенность позволяет им быть спонтанно-активными и направлять в нервные центры разнообразную информацию о сокращении, расслаблении, растяжении, смещении висцеральных органов. Медленноадаптирующиеся рецепторы связаны с тонкими миелинизированными и безмиелиновыми нервными волокнами.
Хеморецепторы активируются при изменении химического состава ткани, например напряжения СО2 и О2 в крови. В органах пищеварения выделены специальные кислото- и щелочечувствительные рецепторы, чувствительные к действию только аминокислот или аминокислот и глюкозы.
Тепловые и холодовые терморецепторы также обнаружены по преимуществу в пищеварительном тракте. Осморецепторы, ионорецепторы (например, натриевые) висцеральных органов обнаружены в печени. Частота их разрядов находится в прямой зависимости от осмотического давления жидкости. Существование специфических ноцицепторов пока еще окончательно не установлено, хотя их роль и отводится некоторым свободным нервным окончаниям. Болевые ощущения возникают при чрезмерной стимуляции любого типа — растяжении, сокращении, действии химических стимулов.
Помимо специфических, имеются и рецепторы, воспринимающие раздражение любой модальности, будь то механическое, химическое, термическое, осмотическое. Местом локализации таких полимодальных интероцепторов является, например, слизистая оболочка пищеварительного тракта.
Все рассмотренные виды висцеральной чувствительности передаются в центры по волокнам трех основных нервных путей: блуждающего, чревных (большого, малого, поясничных) и тазового (рис. 4.23). Из них самым мощным коллектором висцеральной чувствительности является блуждающий нерв. Соотношение в нем афферентных и эфферентных волокон составляет 9:1, в то время как в чревном и тазовом нервах 3:1 и 1:1 соответственно.
Помимо местных сетей, афферентные сигналы могут запускать центральные нейронные механизмы ряда систем: сегментарную, межсегментарную, проприоспинальную, надсегментарную. Несмотря на такую сложность многоступенчатой организации, основа механизма взаимодействия на всех ступенях остается одной и той же: это синаптическая конвергенция к центральным клеткам сигналов различной природы (висцеральной и соматической) и разной модальности, оценка их аппаратом суммации постсинаптических потенциалов и на основании оценки результатов — генерация нового сигнала. Различия между механизмами, включающимися на разных уровнях, непринципиальны и заключаются в количественных особенностях конвергенции. Эти механизмы вовлекаются в разной степени в зависимости от интенсивности воздействия и включаются в разных соотношениях.
Для запуска менее сложных сегментарных механизмов оказывается достаточной и менее сложная суммация постсинаптических процессов. Эти механизмы срабатывают при возбуждении небольшого числа афферентов. Включение более сложных систем требует значительной суммации процессов, а стало быть, более интенсивного притока афферентации. Следовательно, для запуска различных нейронных систем основой является мощность поступающего афферентного потока.
Импульсы, интегрированные в общей системе вставочных нейронов, способны вызвать их активность и, как следствие, появление вегетативных, например дыхательных или сердечно-сосудистых, эффектов. Импульсы могут также активировать клетки высших отделов центральной нервной системы, вызывая появление поведенческих реакций и субъективных ощущений.
Реакция на афферентный импульс и элементы его переработки на подкорковом уровне является основой для последующих процессов в коре больших полушарий, направленных на регуляцию функций определенной висцеральной системы — пищеварительной, дыхательной и т. д. Эти процессы выражаются в виде вызванных потенциалов — первичного и вторичного ответов: первичные сравниваются с пусковыми, вторичные — с корригирующими влияниями коры большого мозга.
При анализе локализации представительства висцеральных систем в коре большого мозга обнаруживается несоответствие числа зон проекций блуждающего и чревного нервов. Объясняется это тем, что блуждающий нерв по числу сенсорных волокон и особенно по величине иннервируемых областей не имеет себе равных, охватывая большое количество внутренних органов, некоторые из которых подвержены в какой-то мере произвольному контролю.
Представительства функционально близких висцеральных систем находятся и в близко расположенных областях коры. Например, зоны брыжеечных, селезеночных и чревных нервов перекрываются представительством блуждающего нерва, что служит основой для тонкой координации процессов, осуществляемых корой больших полушарий, восстановления функции, надежности работы висцеральных органов.
Предложенная В. Н. Черниговским схема проведения висцеральных сигналов в центральной нервной системе дает представление об участии той или иной наиболее важной структуры в этом процессе, хотя и не указывает на степень участия каждой и не отражает всей сложности существующих взаимодействий (схема 4.2). Сигналы, вызывающие ответы в клетках коры большого мозга, после соответствующей обработки передаются в специальные выходы передних отделов поясной извилины, и уже оттуда через гипоталамус нисходящие пути следуют к вставочным (преганглионарным), затем к эффекторным нейронам и далее к исполнительным органам. Таким образом, информация от высших центров по нисходящим путям и от периферических висцеральных и соматических клеток по спинальным дугам поступает к преганглионарным нейронам.
Тело преганглионарного автономного нейрона располагается в сером веществе в одних случаях ствола мозга, в других — спинного мозга. На периферии за пределами спинного мозга нервное волокно вступает в синаптический контакт с эффекторным нейроном. Исключение составляет лишь часть волокон, следующих в составе чревного нерва к надпочечнику. Эти волокна проникают непосредственно в мозговой слой железы, который и выполняет своеобразную функцию постганглионарного звена рефлекторной дуги. Истинное же эффекторное звено дуги автономного рефлекса представляет собой нервную клетку, мигрировавшую из ЦНС.
Преганглионарные волокна различаются по своим функциональным свойствам. Наибольшее их число составляют тонкие, легко возбудимые, с медленным проведением возбуждения единицы. Приближаясь к эффекторным нейронам, преганглионарные волокна теряют миелин и разветвляются на тонкие терминалы, образуя на теле и отростках эффекторного нейрона синаптические контакты.
Эффекторных нейронов несравненно больше, чем преганглионарных волокон. Например, в верхнем шейном симпатическом ганглии одно преганглионарное симпатическое волокно контактирует более чем с сотней эффекторных нейронов. При этом на одном и том же эффекторном нейроне могут оканчиваться разветвления нескольких преганглионарных волокон. Наличие таких широких конвергентных и дивергентных отношений обеспечивает надежность проведения возбуждения. Эта закономерность касается только симпатической части автономной нервной системы, в двух других ее частях подобная конвергенция практически отсутствует.
В интеграции сигналов в низших центрах вегетативной периферии значительная роль отводится пространственной и временной суммации постсинаптических потенциалов. Роль их состоит в том, что поступающие по пресинаптическим терминалям присущие им относительно слабые сигналы благодаря этим процессам трансформируются, превращаясь в сверхпороговые постсинаптические потенциалы эфферентного нейрона.
Тело эффекторной клетки дуги автономного рефлекса представляет собой мигрировавшую из спинного мозга клетку, располагающуюся в одном из периферических автономных ганглиев (рис. 4.24). Нейроны этих ганглиев охватывают своим влиянием, как правило, большие территории висцеральных органов. Ганглии могут располагаться либо около позвоночника (превертебральные), либо в сплетениях вблизи внутренних органов (паравертебральные), наконец, в тканях внутренних органов (интрамуральные, интервисцеральные).
Эффекторный нейрон дуги автономного рефлекса по электрическим показателям в покоящемся состоянии мало чем отличается от мотонейрона соматической дуги. Однако кратковременная или одиночная стимуляция преганглионарных волокон вызывает появление в нем сложной последовательности медленных деполяризующих и гиперполяризующих постсинаптических процессов. В этом случае вначале возникает локальный отрицательный потенциал (О-волна), переходящий в положительную П-волну. Последняя сменяется поздней отрицательной П-волной. Каждая из этих фаз отражает межнейронную передачу, при этом О-волна — возникающий в холинергических синапсах возбуждающий постсинаптический потенциал (ВПСП). Появление П-волны обусловлено возбуждением особой группы преганглионарных волокон, которые оканчиваются на хромаффинных клетках ганглия. Появление деполяризующихся и гиперполяризующихся постсинаптических потенциалов опосредуется мускариноподобным действием ацетилхолина, в то время как гиперполяризация — специальными вставочными адренергическими клетками, регулируя тем самым возбудимость эффекторных нейронов. Как правило, эффекторный нейрон может иметь, помимо основного возбуждающего холинергического преганглионарного входа, еще и прямой вход сугубо периферического происхождения, представляющий одно из звеньев местной рефлекторной дуги ганглионарного уровня. Электрофизиологические характеристики эффекторного нейрона позволяют интегрировать эти сигналы и формировать новый выходной сигнал. Благодаря этим местным дугам в эфферентном нейроне поддерживается необходимый уровень спонтанной активности и при децентрализации ганглия сохраняется его рефлекторная функция. У спонтанно активных эфферентных нейронов фоновые разряды характеризуются низкой частотой. Они могут возникать синхронно пульсовым толчкам, дыхательным и перистальтическим движениям. Паттерн и ритм разрядов совпадают с показателями преганглионарных волокон или активностью волокон местных рефлекторных дуг.
Симпатическая часть
Симпатическая часть автономной нервной системы (рис. 4.25) имеет центральный аппарат, или спинномозговой (торако-люмбальный) центр Якобсона, который представлен симпатическим ядром бокового рога серого вещества спинного мозга. Это ядро простирается от I—II грудных до II—IV поясничных сегментов. Отростки составляющих ядро клеток называются преганглионарными волокнами. Они выходят из спинного мозга в составе его передних корешков через межпозвоночные отверстия. Вскоре после выхода симпатические волокна отделяются от двигательных соматических (см. рис. 4.15) и далее в виде белых соединительных ветвей вступают в узлы пограничного симпатического ствола. Часть волокон образует здесь синаптические контакты с клетками узлов, часть проходит узлы транзитом и вступает в синаптический контакт либо с клетками других узлов пограничного симпатического ствола, либо превертебральных (чревное сплетение, нижнее брыжеечное сплетение) узлов.
Периферический отдел симпатической части автономной нервной системы образован эфферентными и чувствительными нейронами и их отростками, располагающимися в удаленных от спинного мозга узлах. В околопозвоночных, или паравертебральных, узлах часть преганглионарных симпатических волокон синаптически оканчивается на эфферентных нейронах. Волокна эфферентных нейронов, именуемые постганглионарными, разделяются на две группы. Волокна одной из них в виде серых соединительных ветвей вновь вступают в соматический нерв и в его составе без перерыва достигают эффекторного органа (сосуды кожи, мышц), волокна другой группы, собравшись в отдельные веточки, образуют обособленный стволик, направляющийся либо непосредственно к исполнительным органам, либо к предпозвоночным узлам, а через них далее также к исполнительным органам. Постганглионарные волокна в большинстве своем лишены миелиновой оболочки, поэтому имеют розово-серую окраску. Серые ветви отходят от всех узлов пограничного симпатического ствола, который делится на шейную, грудную, поясничную, крестцовую части.
Предпозвоночные, или превертебральные, узлы лежат на большом расстоянии от центральной нервной системы. На их эффекторных нейронах заканчиваются прошедшие, не прерываясь через узлы пограничного симпатического ствола, преганглионарные волокна.
Основную массу узлов составляют нервные клетки. В строме ганглиев найдены чувствительные окончания. В синапсах отчетливо выделяются пре- и постсинаптические мембраны, отмечается большое количество пузырьков, митохондрий, трубочек эндоплазматической сети.
Парасимпатическая часть
Парасимпатическая часть автономной нервной системы имеет общую структуру, подобную симпатической части: здесь также выделяют центральные и периферические образования. Как и в симпатической части, передача возбуждения к исполнительному органу осуществляется по двухнейронному пути. Вместе с тем ряд признаков отличает парасимпатическую часть от симпатической.
Во-первых, центральные структуры парасимпатической части расположены в трех различных, далеко отстоящих друг от друга участках мозга; во-вторых, характерно наличие значительно более длинных преганглионарных и чрезвычайно коротких постганглионарных волокон; в-третьих, парасимпатические волокна иннервируют, как правило, только определенные зоны тела, которые также снабжаются симпатической, а в значительной части, кроме того, и метасимпатической иннервацией.
Центральные образования парасимпатической части автономной нервной системы включают ядра, лежащие в среднем, продолговатом и спинном мозге. В среднем мозге находится парасимпатическое добавочное ядро глазодвигательного нерва (ядро Якубовича, Вестфаля — Эдингера), расположенное вблизи передних бугров четверохолмия; в продолговатом мозге — три пары ядер, от которых начинаются преганглионарные волокна, выходящие из мозга в составе VII, IX, X пар черепных нервов (лицевого, языкоглоточного, блуждающего). Здесь проходят слюноотделительные, слезоотделительные, а также двигательный и секреторный пути для внутренних органов (блуждающий нерв). Парасимпатические ядра спинного мозга располагаются в области I—III или II—IV крестцовых сегментов в боковых рогах серого вещества.
Периферические структуры парасимпатической части автономной нервной системы включают нервные волокна и соответствующие ганглии. Преганглионарные волокна из среднего мозга выходят сбоку от ножек большого мозга в составе глазодвигательного нерва, проникают через глазную щель в глазницу и синаптически заканчиваются на эффекторных клетках расположенного в глубине глазницы ресничного узла. От него отходят два коротких ресничных нерва. Составляющие их постганглионарные волокна вступают в глазное яблоко, разветвляясь в аккомодационной мышце и сфинктере зрачка.
В продолговатом мозге нервные волокна из верхнего слюноотделительного ядра идут в составе лицевого нерва и, покидая его, образуют барабанную струну, которая позже присоединяется к язычному нерву. Последний достигает челюстного или подъязычного узла, постганглионарные волокна которого иннервируют подчелюстную слюнную железу. Преганглионарные волокна, выходящие из нижнего слюноотделительного ядра, вступают в языкоглоточный нерв и далее попадают в ушной узел. Его постганглионарные волокна являются секреторными для околоушной слюнной железы. Преганглионарные волокна из ядер слезоотделительного пути через лицевой нерв вступают в крылонебный узел, постганглионарные волокна которого достигают слюнной железы, желез слизистой оболочки носа и неба.
Блуждающий нерв является смешанным: он включает афферентные и эфферентные парасимпатические, чувствительные и двигательные соматические, а также эфферентные симпатические волокна. По выходе из черепа нерв образует два последовательно лежащих узла: верхний и нижний (яремный и узловой). Верхний узел содержит в основном чувствительные клетки, аналогичные клеткам спинномозговых узлов. От нижнего узла берут начало сердечный депрессорный нерв, возвратный гортанный нерв, пищеводные ветви. У корня легкого от блуждающего нерва отходят соответствующие веточки к легкому. В брюшной полости нерв переходит на желудок, формируя желудочное сплетение, от которого отходят стволики в чревное (солнечное) сплетение. Грудная и брюшная части блуждающего нерва могут рассматриваться лишь как проводники, связывающие центральные структуры с эффекторным аппаратом метасимпатической нервной системы.
Крестцовый отдел парасимпатической части нервной системы представлен тазовым нервом, который направляется к поверхности прямой кишки, где вместе с подчревным симпатическим нервом участвует в образовании тазового сплетения.
Метасимпатическая часть
Структура метасимпатической части отличается относительной простотой (см. рис. 4.18). Здесь нет ядерных образований и система представлена лишь комплексом интрамуральных ганглионарных структур, залегающих в стенках полых висцеральных органов. В соответствии с иннервационными территориями в ней различают энтеральную, кардиальную, респираторную и другие области. Метасимпатическая часть обладает многими признаками, которые отличают ее от других частей автономной нервной системы. Прежде всего эта часть иннервирует только внутренние органы, наделенные моторным ритмом. В сфере ее управления находятся гладкие мышцы, всасывающий и секретирующий эпителий, локальный кровоток, местные эндокринные и иммунные элементы. Метасимпатическая часть получает внешние синаптические входы от симпатической и парасимпатической частей автономной нервной системы и не имеет прямых синаптических контактов с эфферентной частью соматической рефлекторной дуги. Метасимпатическая часть характеризуется наличием собственного сенсорного звена. Представляя базовую иннервацию висцеральных органов, она обладает гораздо большей, чем симпатическая и парасимпатическая части автономной нервной системы, независимостью от ЦНС.
Органы с разрушенными или выключенными с помощью ганглиоблокаторов метасимпатическими путями утрачивают присущую им способность к координированной моторной деятельности и другим функциональным отправлениям.
Особенности конструкции автономной нервной системы
Первое и основное отличие строения автономной нервной системы от строения соматической состоит в расположении эфферентного (моторного) нейрона (см. рис. 4.22). В соматической нервной системе вставочный и моторный нейроны располагаются в сером веществе спинного мозга, в автономной нервной системе эффекторный нейрон вынесен на периферию, за пределы спинного мозга, и лежит в одном из ганглиев — пара-, превертебральном или интраорганном. Более того, в метасимпатической части автономной нервной системы весь рефлекторный аппарат полностью находится в интрамуральных ганглиях и нервных сплетениях внутренних органов.
Второе отличие касается выхода нервных волокон из ЦНС. Соматические нервные волокна покидают спинной мозг сегментарно и перекрывают иннервацией не менее трех смежных сегментов. Волокна же автономной нервной системы выходят из трех участков ЦНС — головного мозга, грудопоясничного и крестцового отделов спинного мозга. Они иннервируют все органы и ткани без исключения. Большинство висцеральных систем имеет тройную — симпатическую, парасимпатическую и метасимпатическую — иннервацию.
Третье отличие касается иннервации органов соматической и автономной нервной системой. Перерезка у животных вентральных корешков спинного мозга сопровождается полным перерождением всех соматических эфферентных волокон. Она не затрагивает дуги автономного рефлекса ввиду того, что ее эффекторный нейрон вынесен в пара- или превертебральный ганглий. В этих условиях эффекторный орган управляется импульсами данного нейрона. Именно это обстоятельство подчеркивает относительную автономию указанного отдела нервной системы.
Четвертое отличие относится к свойствам нервных волокон. В автономной нервной системе они в большинстве своем безмякотные или тонкие мякотные, как, например, преганглионарные волокна, диаметр которых не превышает 5 мкм. Такие волокна принадлежат к типу В. Постганглионарные волокна еще тоньше, большая часть их лишена миелиновой оболочки, они относятся к типу С. В отличие от них соматические эфферентные волокна толстые, мякотные, диаметр их составляет 12—14 мкм. Кроме того, пре- и постганглионарные волокна отличаются низкой возбудимостью. Для вызова в них ответной реакции необходима значительно большая, чем для моторных соматических волокон, сила раздражения. Волокна автономной нервной системы характеризуются большим рефрактерным периодом и большой хронаксией (1,0—2,0 и 0,1—0,8 сигмы соответственно). Скорость распространения по ним нервных импульсов невелика и составляет в преганглионарных волокнах до 18 м/с, в постганглионарных — до 3 м/с. Потенциалы действия волокон автономной нервной системы характеризуются большей, чем в соматических эфферентах, длительностью. Их возникновение в преганглионарных волокнах сопровождается продолжительным следовым положительным потенциалом, в постганглионарных волокнах — следовым отрицательным потенциалом с последующей продолжительной следовой гиперполяризацией (300—400 мс).
Автономный (вегетативный) тонус
В естественных условиях симпатические и парасимпатические центры, а также эффекторные нейроны метасимпатической части автономной нервной системы находятся в состоянии непрерывного возбуждения, получившего название «тонус». Характерной особенностью тонического влияния является длительное поддержание внешнего эффекта, который наиболее выраженно отражается на функциональном состоянии сосудистой стенки, сердечной мышцы, висцеральных органов в целом.
Тоническое состояние можно непосредственно зарегистрировать в отдельных волокнах и клетках всех трех частей автономной нервной системы и оценить по показателям их активности. Частота тонических разрядов в пре- и постганглионарных симпатических волокнах составляет от 0,1 до 5,0 имп/с и находится в зависимости от иннервируемых гладкомышечных органов, которые имеют в свою очередь и собственный базальный мышечный тонус. Тонус можно оценить еще и косвенно. В этом случае основным показателем является изменение деятельности органа после перерезки или электрической стимуляции иннервирующих его волокон. Иллюстрацией этого могут служить классические опыты с одновременной перерезкой на шее собаки обоих блуждающих нервов и односторонней перерезкой на шее кролика симпатического нерва. Перерезка блуждающих нервов вызывает отчетливое учащение сердечного ритма. Перерезка шейного симпатического ствола сопровождается немедленным расширением сосудов уха на стороне перерезки, что является результатом исключения возбуждащего сосудосуживающего влияния. Стимуляция периферических концов перерезанных нервов с частотой 1—2 имп/с приводит к восстановлению исходного сердечного ритма в опытах с перерезкой блуждающих нервов и полному возвращению к уровню сужения сосудов уха, который был до перерезки симпатического ствола.
Преобладание парасимпатического тонуса обычно оценивается на основании частоты сердечных сокращений. Тоническая импульсация, следующая из центров продолговатого мозга по волокнам блуждающего нерва, оказывает на сердце отрицательное хронотропное действие, снижая частоту сердечных сокращений. Напротив, ослабление тонуса ведет к учащению сердечного ритма.
Исключительна роль симпатической части автономной нервной системы и в создании общего сосудистого тонуса. Тонические влияния из сосудодвигательного центра приспосабливают сосуды мелкого и среднего диаметра к местным и общим потребностям организма. В своих тонических влияниях симпатическая часть автономной нервной системы часто взаимодействует с мозговым веществом надпочечников. В этом случае сосудосуживающие реакции усиливаются выбросом адреналина, возникающим в результате активации надпочечников под действием импульсов из сосудодвигательного центра.
Преобладание тонических влияний парасимпатической и симпатической частей автономной нервной системы послужило основанием для создания конституционной классификации. Согласно этой классификации, преобладание в организме тонуса парасимпатической части автономной нервной системы именуется ваготонией, симпатической — симпатикотонией. Ваготония характеризуется замедленным пульсом, склонностью к покраснениям, потливостью, желудочными расстройствами. Для симпатикотонии, напротив, типичным является учащенный пульс и т. д. Чистые формы ваготонии и симпатикотонии встречаются исключительно редко.
Многие стороны природы тонической активности остаются малоизвестными. Считают, что тонус ядерных образований формируется преимущественно благодаря притоку сенсорной информации из рефлексогенных зон, отдельных групп интероцепторов, а также соматических рецепторов. При этом не исключается и существование собственных водителей ритма — пейсмекеров, локализованных в основном в продолговатом мозге. В пользу такой точки зрения свидетельствует возникновение тахикардии после денервации каротидного синуса (sinus caroticus) или области дуги аорты, а также исчезновение разрядов в сердечных веточках блуждающего нерва при снижении артериального давления. Особенностью метасимпатической части автономной нервной системы является существование в ее функциональных модулях специальных клеток-осцилляторов, так называемых водителей ритма (рис. 4.26). Эти клетки не имеют синаптических входов, на их функцию не влияют ганглиоблокаторы и вещества медиаторного типа, однако сами они синаптически связаны со вставочными и эффекторными нейронами. Спонтанная деполяризация этих водителей ритма создает и постоянно поддерживает необходимый уровень тонической активности.
В целом тонус автономной нервной системы рассматривается как одно из проявлений гомеостатического состояния и одновременно один из механизмов его стабилизации.
Синаптическая передача возбуждения в автономной нервной системе
У позвоночных животных в автономной нервной системе имеется три вида синаптической передачи: электрическая, химическая и смешанная. Органом с типичными электрическими синапсами является цилиарный ганглий птиц, лежащий в глубине глазницы у основания глазного яблока. Передача возбуждения здесь осуществляется практически без задержки в обоих направлениях. К редко встречающимся можно отнести и передачу через смешанные синапсы, в которых одновременно соседствуют структуры электрических и химических синапсов. Этот вид также характерен для цилиарного ганглия птиц. Основным же способом передачи возбуждения в автономной нервной системе является химический. Он осуществляется по определенным закономерностям, среди которых выделяют два принципа. Первый (принцип Дейла) заключается в том, что нейрон со всеми отростками выделяет один медиатор. Как стало теперь известно, наряду с основным в этом нейроне могут присутствовать также другие передатчики и участвующие в их синтезе вещества. Согласно второму принципу, действие каждого медиатора на нейрон или эффектор зависит от природы рецептора постсинаптической мембраны.
В автономной нервной системе насчитывают более десяти видов нервных клеток, которые продуцируют в качестве основных разные медиаторы: ацетилхолин, норадреналин, серотонин и другие биогенные амины, аминокислоты, АТФ. В зависимости от того, какой основной медиатор выделяется окончаниями аксонов автономных нейронов, эти клетки принято называть холинергическими, адренергическими, серотоиинергическими, пуринергическими и т. д. нейронами.
Каждый из медиаторов выполняет передаточную функцию, как правило, в определенных звеньях дуги автономного рефлекса (рис. 4.27). Так, ацетилхолин выделяется в окончаниях всех преганглионарных симпатических и парасимпатических нейронов, а также большинства постганглионарных парасимпатических окончаний. Кроме того, часть постганглионарных симпатических волокон, иннервирующих потовые железы и, по-видимому, вазодилататоры скелетных мышц, также осуществляют передачу с помощью ацетилхолина. В свою очередь норадреналин является медиатором в постганглионарных симпатических окончаниях (за исключением нервов потовых желез и симпатических вазодилататоров) — сосудов сердца, печени, селезенки.
Медиатор, освобождающийся в пресинаптических терминалах под влиянием приходящих нервных импульсов, взаимодействует со специфическим белком-рецептором постсинаптической мембраны и образует с ним комплексное соединение. Белок, с которым взаимодействует ацетилхолин, носит название холинорецептора, адреналин или норадреналин — адренорецептора и т. д. Местом локализации рецепторов различных медиаторов является не только постсинаптическая мембрана. Обнаружено существование и специальных пресинаптических рецепторов, которые участвуют в механизме обратной связи регуляции медиаторного процесса в синапсе.
Помимо холино-, адрено-, пуринорецепторов, в периферической части автономной нервной системы имеются рецепторы пептидов, дофамина, простагландинов. Все виды рецепторов, вначале обнаруженные в периферической части автономной нервной системы, были найдены затем в пре- и постсинаптических мембранах ядерных структур ЦНС.
Характерной реакцией автономной нервной системы является резкое повышение ее чувствительности к медиаторам после денервации органов. Например, после ваготомии орган обладает повышенной чувствительностью к ацетилхолину, соответственно после симпатэктомии — к норадреналину. Полагают, что в основе этого явления лежит резкое возрастание числа соответствующих рецепторов постсинаптической мембраны, а также снижение содержания или активности ферментов, расщепляющих медиатор (ацетилхолин-эстераза, моноаминоксидаза и др.).
В автономной нервной системе, помимо обычных эффекторных нейронов, существуют еще специальные клетки, соответствующиепостганглионарным структурам и выполняющие их функцию. Передача возбуждения к ним осуществляется обычным химическим путем, а отвечают они эндокринным способом. Эти клетки получили название трансдукторов. Их аксоны не формируют синаптических контактов с эффекторными органами, а свободно заканчиваются вокруг сосудов, с которыми образуют так называемые гемальные органы. К трансдукторам относят следующие клетки: 1) хромаффинные клетки мозгового слоя надпочечников, которые на холинергический передатчик преганглионарного симпатического окончания отвечают выделением адреналина и норадреналина; 2) юкста-гломерулярные клетки почки, которые отвечают на адренергический передатчик постганглионарного симпатического волокна выделением в кровяное русло ренина; 3) нейроны гипоталамических супраоптического и паравентрикулярного ядер, реагирующие на синаптический приток разной природы выделением вазопрессина и окситоцина; 4) нейроны ядер гипоталамуса.
Действие основных классических меадиаторов может быть воспроизведено с помощью фармакологических препаратов. Например, никотин вызывает эффект, подобный эффекту ацетилхолина, при действии на постсинаптическую мембрану постганглионарного нейрона, в то время как сложные эфиры холина и токсин мухомора мускарин — на постсинаптическую мембрану эффекторной клетки висцерального органа. Следовательно, никотин вмешивается в межнейронную передачу в автономном ганглии, мускарин — в нейро-эффекторную передачу в исполнительном органе. На этом основании считают, что имеется соответственно два типа холинорецепторов: никотиновые (Н-холинорецепторы) и мускариновые (М-холинорецепторы). В зависимости от чувствительности к различным катехоламинам адренорецепторы делят на α-адренорецепторы и β-адренорецепторы. Их существование установлено посредством фармакологических препаратов, избирательно действующих на определенный вид адренорецепторов.
В ряде висцеральных органов, реагирующих на катехоламины, находятся оба вида адренорецепторов, но результаты их возбуждения бывают, как правило, противоположными (табл. 4.2). Например, в кровеносных сосудах скелетных мышц имеются α- и β-адренорецепторы. Возбуждение α-адренорецепторов приводит к сужению, а β-адренорецепторов — к расширению артериол. Оба вида адренорецепторов обнаружены и в стенке кишки, однако реакция органа при возбуждении каждого из видов будет однозначно характеризоваться торможением активности гладких мышечных клеток. В сердце и бронхах нет α-адренорецепторов и медиатор взаимодействует только с β-адренорецепторами, что сопровождается усилением сердечных сокращений и расширением бронхов. В связи с тем что норадреналин вызывает наибольшее возбуждение β-адренорецепторов сердечной мышцы и слабую реакцию бронхов, трахеи, сосудов, первые стали называть β1-адренорецепторами, вторые — β2-адренорецепторами.
При действии на мембрану гладкой мышечной клетки адреналин и норадреналин активируют находящуюся в клеточной мембране аденилатциклазу. При наличии ионов Mg2+ этот фермент катализирует образование в клетке цАМФ (циклического 3' ,5' -аденозинмонофосфата) из АТФ. Последний продукт в свою очередь вызывает ряд физиологических эффектов, активируя энергетический обмен, стимулируя сердечную деятельность.
Особенностью адренергического нейрона является то, что он обладает чрезвычайно длинными тонкими аксонами, которые разветвляются в органах и образуют густые сплетения. Общая длина таких аксонных терминалей может достигать 30 см. По ходу терминалей имеются многочисленные расширения — варикозы, в которых синтезируется, запасается и выделяется медиатор. С приходом импульса норадреналин одновременно выделяется из многочисленных расширений, действуя сразу на большую площадь гладкомышечной ткани. Таким образом, деполяризация мышечных клеток сопровождается одновременным сокращением всего органа.
Различные лекарственные средства, оказывающие на эффекторный орган действие, аналогичное действию постганглионарного волокна (симпатического, парасимпатического и т.п.), получили название миметиков (адрено-, холиномиметики). Наряду с этим имеются и вещества, избирательно блокирующие функцию рецепторов постсинаптической мембраны. Они названы ганглиоблокаторами. Например, аммониевые соединения избирательно выключают Н-холинорецепторы, а атропин и скополамин — М-холинорецепторы.
Классические медиаторы выполняют не только функцию передатчиков возбуждения, но обладают и общебиологическим действием. К ацетилхолину наиболее чувствительна сердечно-сосудистая система, он вызывает и усиленную моторику пищеварительного тракта, активируя одновременно деятельность пищеварительных желез, сокращает мускулатуру бронхов и понижает бронхиальную секрецию. Под влиянием норадреналина происходит повышение систолического и диастолического давления без изменения сердечного ритма, усиливаются сердечные сокращения, снижается секреция желудка и кишки, расслабляется гладкая мускулатура кишки и т. д. Более разнообразным диапазоном действий характеризуется адреналин. Посредством одновременной стимуляции ино-, хроно- и дромотропной функций адреналин повышает сердечный выброс. Адреналин оказывает расширяющее и антиспазматическое действие на мускулатуру бронхов, тормозит моторику пищеварительного тракта, расслабляет стенки органов, но тормозит деятельность сфинктеров, секрецию желез пищеварительного тракта.
В тканях всех видов животных обнаружен серотонин (5-окситриптамин). В мозге он содержится преимущественно в структурах, имеющих отношение к регуляции висцеральных функций, на периферии продуцируется энтерохромаффинными клетками кишки. Серотонин является одним из основных медиаторов метасимпатической части автономной нервной системы, участвующей преимущественно в нейроэффекторной передаче, и выполняет также медиаториую функцию в центральных образованиях. Известно три типа серотонинергических рецепторов — Д, М, Т. Рецепторы Д-типа локализованы в основном в гладких мышцах и блокируются диэтиламидом лизергиновой кислоты. Взаимодействие серотонина с этими рецепторами сопровождается мышечным сокращением. Рецепторы М-типа характерны для большинства автономных ганглиев; блокируются морфином. Связываясь с этими рецепторами, передатчик вызывает ганглиостимулирующий эффект. Рецепторы Т-типа, обнаруженные в сердечной и легочной рефлексогенных зонах, блокируются тиопендолом. Действуя на эти рецепторы, серотонин участвует в осуществлении коронарных и легочных хеморефлексов. Серотонин способен оказывать прямое действие на гладкую мускулатуру. В сосудистой системе оно проявляется в виде констрикторных или дилататорных реакций. При прямом действии сокращается мускулатура бронхов, при рефлекторном — изменяются дыхательный ритм и легочная вентиляция. Особенно чувствительна к серотонину пищеварительная система. На введение серотонина она реагирует начальной спастической реакцией, переходящей в ритмические сокращения с повышенным тонусом и завершающейся торможением активности.
Для многих висцеральных органов характерной является пуринергическая передача, названная так вследствие того, что при стимуляции пресинаптических терминален выделяются аденозин и инозин — пуриновые продукты распада. Медиатором же в этом случае является А Т Ф. Местом его локализации служат пресинаптические терминалы эффекторных нейронов метасимпатической части автономной нервной системы.
Выделившийся в синаптическую щель АТФ взаимодействует с пуринорецепторами постсинаптической мембраны двух типов. Пуринорецепторы первого типа более чувствительны к аденозину, второго — к АТФ. Действие медиатора направлено преимущественно на гладкую мускулатуру и проявляется в виде ее релаксации. В механизме кишечной пропульсии пуринергические нейроны являются главной антагонистической тормозной системой по отношению к возбуждающей холинергической системе. Пуринергические нейроны участвуют в осуществлении нисходящего торможения, в механизме рецептивной релаксин желудка, расслабления пищеводного и анального сфинктеров. Сокращения кишечника, возникающие вслед за пуринергически вызванным расслаблением, обеспечивают соответствующий механизм прохождения пищевого комка.
В числе медиаторов может быть гистамин. Он широко распространен в различных органах и тканях, особенно в пищеварительном тракте, легких, коже. Среди структур автономной нервной системы наибольшее количество гистамина содержится в постганглионарных симпатических волокнах. На основании ответных реакций в некоторых тканях обнаружены и специфические гистаминовые (Н-рецепторы) рецепторы: Н1- и Н2-рецепторы. Классическим действием гистамина является повышение капиллярной проницаемости и сокращение гладкой мускулатуры. В свободном состоянии гистамин снижает кровяное давление, уменьшает частоту сердечных сокращений, стимулирует симпатические ганглии.
На межнейронную передачу возбуждения в ганглиях автономной нервной системы тормозное влияние оказывает ГАМК. Как медиатор она может принимать участие в возникновении пресинаптического торможения.
Большие концентрации различных пептидов, особенно субстанции Р, в тканях пищеварительного тракта, гипоталамуса, задних корешков спинного мозга, а также эффекты стимуляции последних и другие показатели послужили основанием считать субстанцию Р медиатором чувствительных нервных клеток.
Помимо классических медиаторов и «кандидатов» в медиаторы, в регуляции деятельности исполнительных органов участвует еще большое число биологически активных веществ — местных гормонов. Они регулируют тонус, оказывают корригирующее влияние на деятельность автономной нервной системы, им принадлежит существенная роль в координации нейрогуморальной передачи, в механизмах выделения и действия медиаторов.
В комплексе активных факторов видное место занимают простагландины, которых много содержится в волокнах блуждающего нерва. Отсюда они выделяются спонтанно либо под влиянием стимуляции. Существует несколько классов простагландинов: Е, G, А, В. Их основное действие — возбуждение гладких мышц, угнетение желудочной секреции, релаксация мускулатуры бронхов. На сердечно-сосудистую систему они оказывают разнонаправленное действие: простагландины класса А и Е вызывают вазодилатацию и гипотензию, класса G — вазоконстрикцию и гипертензию.
Влияние автономной нервной системы на функции тканей и органов
Главной функцией автономной нервной системы является регулирование процессов жизнедеятельности органов тела, согласование и приспособление их работы к общим нуждам и потребностям организма в условиях окружающей среды. Выражением этой функции служит регуляция метаболизма, возбудимости и других сторон деятельности органов и самой ЦНС. В этом случае управление работой тканей, органов и систем осуществляется посредством двух типов влияний — пусковых и корригирующих.
Пусковые влияния используются в случае, если работа исполнительного органа не является постоянной, а возникает лишь с приходом к нему импульсов по волокнам автономной нервной системы. Если же орган обладает автоматизмом и его функция осуществляется непрерывно, то автономная нервная система посредством своих влияний может усиливать или ослаблять его деятельность в зависимости от потребности. Это будут корригирующие влияния. Пусковые влияния могут дополняться корригирующими.
Влияние автономной нервной системы на висцеральные функции. Все структуры и системы организма иннервируются волокнами автономной нервной системы. Многие из них имеют двойную, а полые висцеральные органы даже тройную (симпатическую, парасимпатическую и метасимпатическую) иннервацию. Изучение роли каждой из них обычно осуществляют с помощью электрического раздражения, хирургического или фармакологического выключения, химической стимуляции и т. д.
Так, сильное раздражение симпатических волокон вызывает учащение сердечных сокращений, увеличение силы сокращения сердца, расслабление мускулатуры бронхов, снижение моторной активности желудка и кишечника, расслабление желчного пузыря, сокращение сфинктеров и другие эффекты (рис. 4.28). Раздражение блуждающего нерва характеризуется противоположным действием: уменьшается ритм и сила сердечных сокращений, расширяются сосуды языка, слюнных желез, половых органов, суживаются бронхи, активизируется работа желудочных желез, расслабляются сфинктеры мочевого пузыря и сокращается его мускулатура (см. табл. 4.2).
Эти наблюдения послужили основанием для представления о существовании «антагонистических» отношений между симпатической и парасимпатической частями автономной нервной системы. Их взаимоотношение уподоблялось коромыслу весов, в которых подъем на определенный уровень одной чаши сопровождается снижением на такой же уровень другой.
Представлению «уравновешивания» симпатических влияний парасимпатическими противоречит ряд фактов: например, слюноотделение стимулируется раздражением волокон симпатической и парасимпатической природы, так что здесь проявляется согласованная реакция, необходимая для пищеварения; ряд органов и тканей снабжается только либо симпатическими, либо парасимпатическими волокнами. К таким органам относятся многие кровеносные сосуды, селезенка, мозговой слой надпочечника, некоторые экзокринные железы, органы чувств и ЦНС.
Известно, что многие внутренние органы, извлеченные из организма, продолжают выполнять присущие им функции. Например, сохраняется перистальтическая и всасывательная функция кишки и т. д. Такая относительная функциональная независимость объясняется наличием в стенках этих органов метасимпатической части автономной нервной системы, которая обладает собственным нейрогенным ритмом, имеет полный набор необходимых для самостоятельной рефлекторной деятельности звеньев — сенсорного, ассоциативного, эффекторного с соответствующим медиаторным обеспечением. В составе этой системы имеются собственные сенсорные элементы (механо-, хемо-, термо-, осморецепторы), которые посылают в свои внутренние сети информацию о состоянии иннервируемого органа, а также способны передавать сигналы в ЦНС. Сфера иннервации метасимпатической части автономной нервной системы ограничена и охватывает сугубо внутренние органы и то не все. Для этих органов метасимпатическая иннервация является базовой, все звенья ее рефлекторного пути локализуются только в интрамуральных ганглиях. Метасимпатическая часть не имеет своего центрального аппарата и ее эфферентные связи с центральными структурами опосредованы нейронами симпатической и парасимпатической частей автономной нервной системы, образующими синаптические контакты на телах и отростках метасимпатических интернейронов и эффекторных нейронов (см. рис. 4.19).
Мнение о том, что метасимпатическая часть автономной нервной системы является диффузным парасимпатическим ганглием, в котором прямые синаптические контакты между преганглионарными волокнами и ганглионарными клетками являются основой для управления (например, сердечной, желудочной или кишечной функции), при экспериментальном рассмотрении не подтверждается. Несостоятельно также и представление о метасимпатической части автономной нервной системы как о третьем нейроне в эфферентном звене симпатического пути. Метасимпатическая часть автономной нервной системы — это относительно независимая самостоятельная интегративная система. Ее функцию можно уподобить микропроцессору, расположенному в непосредственной близости от эффекторов (гладкая мышца, всасывающий и экскретирующий эпителий, экзокринные и эндокринные элементы), которые ею контролируются и регулируются.
Невыгодность размещения в ЦНС аппарата, необходимого для постоянного и непрерывного контроля за каждой из висцеральных функций, подтверждается тем, что только метасимпатическая часть автономной нервной системы в кишечнике имеет такое же число клеток (1*108), что и весь спинной мозг, а число метасимпатических нейронов, приходящихся на 1 см2 поверхности кишечника, составляет около 20 000. Существование специальных местных метасимпатических механизмов регуляции функций имеет определенный физиологический смысл. Их наличие увеличивает надежность регуляции функций. Эта регуляция может происходить в случае выключения связи с центральными структурами. При этом ЦНС освобождается от избыточной информации.
Основная функциональная роль метасимпатической части автономной нервной системы состоит в осуществлении механизмов, обеспечивающих гомеостаз — относительное динамическое постоянство внутренней среды и устойчивость основных физиологических функций. В отличие от нее симпатическая часть автономной нервной системы рассматривается как система тревоги, мобилизации защитных сил и ресурсов для активного взаимодействия с факторами среды. Задачу восстановления и поддержания этого постоянства, нарушенного в результате возбуждения симпатической части автономной нервной системы, берет на себя метасимпатическая и отчасти парасимпатическая части автономной нервной системы.
Автономные (вегетативные) рефлексы. Переключение висцеральных афферентных сигналов на эфферентные клетки может происходить в периферических образованиях автономной нервной системы: пара-, превертебральных и интрамуральных ганглиях, называемых низшими рефлекторными центрами, а также на спинальном уровне. В спинальных структурах имеется специальный интернейронный аппарат, осуществляющий первичную обработку сенсорных сигналов. Этот аппарат соединен с клетками боковых рогов спинного мозга и может согласовывать афферентные сигналы, поступающие одновременно из различных рецептивных зон при раздражении интеро- и экстероцепторов. Интеграция висцеральных и соматических сигналов не ограничивается сегментарным спинномозговым уровнем и в определенных условиях может осуществляться уровнями более высокого порядка (см. рис. 4.17). Их координация осуществляется в центрах, расположенных в ретикулярной формации ствола мозга, мозжечке, гипоталамусе, лимбических образованиях и в коре большого мозга.
Процессы в автономной и соматической нервной системе тесно связаны, хотя в ответ на раздражение висцеральных афферентных волокон автономная и соматическая системы вовлекаются в разной степени. Рефлексы в этом случае разделяются на висцеро-висцеральные, висцеросоматические и висцеросенсорные. Уместно назвать соматовисцеральный рефлекс, а также отметить, что в клинической практике существенное значение отводится еще висцеродермальным и дермовисцеральным рефлексам.
Висцеро-висцеральный рефлекс включает пути, в которых возбуждение возникает и заканчивается во внутренних органах. В этом случае рефлекторные дуги могут быть разного уровня. Одни замыкаются в интрамуральных ганглиях и обеспечиваются метасимпатической иннервацией, другие — в пара- и превертебральных симпатических узлах, наконец, третьи имеют спинальный и более высокий уровень замыкания.
При висцеро-висцеральном рефлексе внутренний орган может отвечать двояко: либо торможением, либо усилением функций.
К числу таких рефлексов относится классический рефлекс Гольца: механическое раздражение брыжейки вызывает замедление частоты сердечных сокращений. Другим примером служит раздражение рецепторов пищеварительного тракта, сопровождающееся ослаблением тонуса мышц, суживающих зрачок. Раздражение каротидной или аортальной рефлексогенных зон влечет за собой изменение интенсивности дыхания, уровня кровяного давления, частоты сердечных сокращений.
Разновидностью висцеро-висцерального является аксон-рефлекс. Это понятие охватывает рефлекторные процессы, осуществляющиеся по разветвлениям аксона без участия тела нервной клетки. Возбуждение возникает в одной ветви аксона, затем переходит на другую и по ней направляется к исполнительному органу, вызывая соответствующую реакцию. Есть и другое объяснение возникновению аксон-рефлекса. Экспериментально доказано, что при возбуждении непосредственно рецепторов из рецепторных мембран выделяются биологически активные вещества типа АТФ и разнообразных пептидов, обладающих вазодилататорным действием, которые вызывают соответствующий эффект.
Понятие аксон-рефлекса используется довольно широко. Им, например, объясняют механизм возникновения сосудистой реакции при раздражении кожных болевых рецепторов. Аксон-рефлекс удается воспроизвести даже после удаления спинного мозга, а также дегенерации симпатических волокон, иннервирующих сосудистую стенку исследуемой области.
Висцеросоматический рефлекс также возникает при раздражении внутренних органов и в дополнение к висцеральным вызывает появление соматических реакций. Они выражаются, например, в изменении текущей активности, сокращении или расслаблении скелетных мышц. Примером такой реакции может служить торможение общей двигательной активности организма при раздражении чувствительных окончаний синокаротидной зоны, а также сокращение мышц брюшной стенки или подергивание конечностей при раздражении рецепторов пищеварительного тракта.
Висцеросенсорный рефлекс осуществляется по тем же путям, что и висцеросоматический, но для его вызова необходимо продолжительное и сильное воздействие. Реакция возникает не только во внутренних органах, соматической мышечной системе, но в дополнение к этому изменяется и соматическая чувствительность. Зона повышенного восприятия обычно ограничивается участком кожи, иннервируемым сегментом, к которому поступают импульсы от раздражаемого висцерального органа. Механизм этого явления основан на том, что висцеральные и кожные чувствительные волокна конвергируют на одних и тех же нейронах спинно-таламического пути, в промежуточных структурах происходит потеря специфичности информации, в результате чего ядерные структуры центральной нервной системы и кора большого мозга связывают возникающее возбуждение с раздражением определенной области кожной поверхности.
Среди рефлексов этого типа особое значение придается висцеродермальному рефлексу, при котором раздражение внутренних органов сопровождается изменением потоотделения, электрического сопротивления (электропроводимости) кожи, изменением кожной чувствительности. Вследствие сегментарной организации автономной и соматической иннервации на ограниченных участках поверхности тела, топография которых различна в зависимости от того, какой орган раздражается, при заболевании внутренних органов возникает повышение тактильной и болевой чувствительности определенных областей кожи. Эти боли названы отраженными, а области их проявления — зонами Захарьина—Геда.
Существует и соматовисцеральный рефлекс, разновидностью которого является дермовисцеральный рефлекс. Он выражается тем, что при раздражении некоторых областей поверхности тела возникают сосудистые реакции и изменения функций определенных висцеральных органов. Это явление послужило основанием для возникновения целого направления клинической медицины — рефлексотерапии.
Адаптационно-трофическая функция симпатической части автономной нервной системы. Л. А. Орбели и сотр. провели исследование функционального значения симпатической иннервации для скелетных мышц, что позволило ему сформулировать учение об адаптационно-трофическом влиянии симпатической части автономной нервной системы. В этом влиянии было выделено два неразрывно связанных компонента: влияния адаптационные и влияния трофические, лежащие в основе адаптационных.
Под адаптационными понимаются влияния симпатической части автономной нервной системы, в результате которых происходит приспособление органов к выполнению тех или иных функциональных нагрузок. Сдвиги наступают благодаря тому, что симпатические влияния оказывают на органы трофическое действие, которое выражается в изменении скорости протекания метаболических процессов.
В 20-х годах А. Г. Гинецинский, изучая влияние симпатических волокон на скелетную мышцу лягушки, обнаружил, что утомленная до полной неспособности сокращаться мышца начинает отвечать на стимуляцию моторных нервов после раздражения ее симпатических волокон вначале слабыми, а потом все более сильными сокращениями (рис. 4.29). Оказалось, что при стимуляции симпатических волокон мышца приобретала способность к развитию более сильного напряжения и более длительного его поддержания даже в условиях тетанического возбуждения. В мышце в этот момент происходят укорочение хронаксии, облегчение перехода возбуждения с нерва на мышцу, повышение чувствительности к ацетилхолину, изменение упруговязких свойств и электрической проводимости, повышение потребления кислорода. В миокарде под влиянием раздражения симпатических волокон возникают изменения в потреблении кислорода, содержания гликогена, креатинфосфата, АТФ, актомиозина, РНК, ДНК, фосфолипидов, гуанин-, аденин-, урацилнуклеотидов в активности ряда ферментов.
Эти влияния распространяются не только на мышечную деятельность, но относятся к работе рецепторов, синапсов, различных отделов ЦНС, эндокринных желез, к протеканию безусловных спинномозговых, вазомоторных и дыхательных рефлексов, а также условнорефлекторной деятельности. Эффекты адаптационно-трофического влияния, полученные сначала при раздражении симпатических волокон, полностью воспроизводятся раздражением гипоталамической области. Следовательно, в целом организме адаптационно-трофические влияния могут осуществляться рефлекторно (посредством стимуляции рецепторов чувствительных путей), а также и путем непосредственного раздражения гипоталамических центров, нейроны которых могут возбуждаться образуемыми местно или приносимыми с кровью биологически активными веществами. Таким образом, адаптационно-трофическое влияние симпатической части автономной нервной системы, не являясь пусковым, модулирует функциональную активность того или иного органа — рецепцию, проведение возбуждения, медиацию, сокращение, секрецию и др. и приспосабливает его к потребностям организма.
Изучение физиологических и биохимических механизмов, лежащих в основе регуляторных влияний симпатической части автономной нервной системы на мышечную ткань, показало, что скелетные мышцы позвоночных животных не имеют специальной симпатической иннервации и ее влияния осуществляются за счет медиаторов — адреналина и норадреналина. Медиаторы достигают моторных пластинок и мышечных волокон путем диффузии. Эти вещества восстанавливают и облегчают нервно-мышечную передачу, увеличивают выделение ацетилхолина волокном двигательного нерва. Медиаторы участвуют также в мобилизации энергетических ресурсов клетки, оказывая влияние на различные пути метаболизма через систему цАМФ, способствуют восстановлению функции утомленной мышцы. Катехоламины могут также увеличивать силу мышечного сокращения путем усиления процессов транспорта кальция внутри клетки.
В различных органах симпатические окончания по-разному расположены по отношению к эффекторным клеткам и другим тканевым элементам. Например, в миокарде одни адренергические окончания подходят непосредственно к эндотелию капилляров или их перицитам, другие — к миоцитам органа, третьи иннервируют одновременно и капилляры, и паренхиматозные клетки, четвертые расположены свободно в межклеточном пространстве. Несмотря на такое разнообразие локализации симпатических окончаний, все клетки во всех тканях испытывают их трофическое влияние. Это связано с тем, что, помимо прямых синаптических контактов, существует еще и несинаптическая доставка медиаторов к клеткам эфферентных органов. Следовательно, адаптационно-трофическое влияние симпатической части автономной нервной системы может быть не только прямым, но и косвенным.
Это подтверждается тем, что, во-первых, в период относительного покоя организма в его жидких средах присутствует значительное количество норадреналина, который попадает в межклеточные пространства, лимфу, цереброспинальную жидкость, кровь из адренергических синапсов и содержание его значительно возрастает при нагрузках и чрезвычайных воздействиях на организм. Во-вторых, адаптационно-трофические влияния осуществляются симпатической частью автономной нервной системы еще и через мозговое вещество надпочечников, которое иннервируется ее преганглионарными волокнами. Это вещество выделяет в кровь адреналин и норадреналин, которые при прямом контакте с органами и тканями вызывают такие же эффекты, как и симпатические окончания. В-третьих, норадреналин и адреналин проникают через гематоэнцефалический барьер в гипоталамическую область. Здесь благодаря наличию специфических рецепторов они воздействуют на передний и задний отделы, аденогипофизарную зону гипоталамуса и включают в процесс эндокринные железы. Гормоны этих желез способны влиять не на все виды метаболизма. В-четвертых, симпатические стимулы, поступающие к органу по нервных волокнам или с кровью, содержащей норадреналин и адреналин, изменяя его трофическое состояние, одновременно изменяют уровень чувствительности органа к гормонам. Следовательно, чувствительность органа к биологически активным веществам является еще одной мерой трофического обеспечения органов и тканей.
Учитывая действие циркулирующих в крови медиаторов на клетки, не соприкасающиеся с нервными окончаниями, и роль клеток, связанных с симпатическими волокнами классическими синапсами, можно представить механизм адаптационно-трофических влияний на клеточные популяции следующим образом. Передатчиками влияния симпатического медиатора являются встроенные в мембрану клеток адренорецепторы, аденилатциклаза, цАМФ, цГМФ. Медиатор активирует эту систему посредством первичного контакта со своим рецептором. Например, норадреналин активирует аденилатциклазу через β-адренорецепторы.
Особое значение в механизме адаптационно-трофического действия отводится в настоящее время нейропептидам, к числу которых относятся фрагменты АКТГ, аналоги вазопрессина и окситоцина, либерины, соматостатин, энкефалины, эндорфины, вещество Р, брадикинин, нейротензин, холецистокинин, их производные и другие пептиды. Эти вещества модулируют действие медиаторов на пресинаптическом и постсинаптическом уровне, влияя на их синтез, выведение, инактивацию. Нейропептиды обладают способностью синтезироваться и проникать в нервную клетку и по ее аксонам перемещаться в пресинаптические терминали. Внутриклеточные эффекты ряда пептидов связаны с аденилатциклазной системой.
Адаптационно-трофическая функция убедительно демонстрируется в опытах с хирургическим, химическим, иммунным удалением симпатической части автономной нервной системы.
Тотальная симпатэктомия в условиях покоя не сопровождается значительными расстройствами висцеральных функций, однако симпатэктомированные животные не могут осуществлять физические усилия, с большим трудом оправляются от кровотечений, шока, гипогликемии, плохо переносят перегревание и охлаждение. У этих животных отсутствует проявление характерных защитных реакций и показателей агрессивности: расширение зрачков, тахикардия, повышение притока крови к скелетным мышцам.
В отличие от симпатической влияния парасимпатической части автономной нервной системы на процессы в организме сравнительно ограничены (см. рис. 4.21). Они могут сказываться либо непосредственно на исполнительных органах, либо через метасимпатическую часть автономной нервной системы. В первом случае постганглионарный нейрон непосредственно контактирует с эффектом и вызываемое им действие зависит главным образом от прямых влияний центральной нервной системы. Во втором случае преганглионарные парасимпатические волокна оканчиваются на интернейроне или мотонейроне функционального модуля метасимпатической части автономной нервной системы, представляющего общий конечный путь для импульсов, поступающих по блуждающему и тазовому нервам (см. рис. 4.18). Здесь они взаимодействуют с импульсами местных метасимпатических сетей.
Центры регуляции висцеральных функций. Координация деятельности всех трех частей автономной нервной системы осуществляется сегментарными и надсегментарными центрами (аппаратами) при участии коры большого мозга (см. рис. 4.7). В сложноорганизованном отделе промежуточного мозга — гипоталамической области, находятся ядра, имеющие непосредственное отношение к регуляции висцеральных функций.
Сегментарные центры вследствие особенностей их организации, закономерностей функционирования и медиации являются истинно автономными. В центральной нервной системе они находятся в спинном мозге и в стволе мозга (отдельные ядра черепных нервов), а на периферии составляют сложную систему из сплетений, ганглиев, волокон.
Надсегментарные центры расположены в головном мозге главным образом на лимбико-ретикулярном уровне. Эти интегративные аппараты мозга обеспечивают целостные формы поведения, адаптацию к меняющимся условиям внешней и внутренней среды. Задачей этих аппаратов является организация деятельности функциональных систем, ответственных за регуляцию психических, соматических и висцеральных функций.
Все эти сложные механизмы регуляции деятельности висцеральных органов и систем условно объединены многоэтажной иерархической структурой. Ее базовым, или первым, структурным уровнем являются внутриорганные рефлексы, замыкающиеся в интрамуральных ганглиях и имеющие метасимпатическую природу. Строго говоря, эти ганглии являются низшими рефлекторными центрами. Второй структурный уровень представлен экстрамуральными паравертебральными ганглиями брыжеечных и чревного сплетений. Оба этих низших этажа обладают отчетливо выраженной автономностью и могут осуществлять регуляцию деятельности висцеральных органов и тканей относительно независимо от центральной нервной системы. Центры спинного мозга и ствола представляют третий структурный уровень. Наконец, гипоталамус, ретикулярная формация, лимбическая система, мозжечок, новая кора венчают пирамиду иерархии (четвертый структурный уровень).
Каждый следующий более высокий уровень регуляции определяет и более высокую степень интеграции висцеральных функций. Например, тонус сосудов отдельных органов или областей тела находится под контролем спинальных симпатических центров, в то время как общий уровень артериального давления находится в компетенции сосудодвигательного центра продолговатого мозга. Что же касается участия в целом сердечно-сосудистой системы в общих реакциях организма, координации взаимодействия висцеральных и соматических систем в сложных поведенческих актах, то они координируются и регулируются высшими этажами нервной системы, т. е. верхушкой условной иерархической пирамиды.
Спинальные центры. В шейной и в начале грудной части (последний шейный, I и II грудные сегменты) располагаются тела преганглионарных симпатических нейронов, иннервирующие гладкие мышцы глазного яблока: мышцу, расширяющую зрачок, глазничную часть круговой мышцы глаза, одну из мышц верхнего века (см. рис. 4.18). Это образование спинного мозга носит название спиноцилиарного центра. Вторые нейроны рассматриваемого эфферентного пути лежат в верхнем шейном симпатическом узле, а их постганглионарные волокна заканчиваются в мышцах глаза. Раздражение центра вызывает расширение зрачка (мидриаз), выпячивание глазного яблока (экзофтальм), раскрытие глазной щели. Разрушение центра или перерезка постганглионарных симпатических волокон вызывает возникновение синдрома Бернара—Горнера — сужение зрачка (миоз), западение глазного яблока (энофтальм), сужение глазной щели.
Пять верхних грудных сегментов служат местом локализации симпатических нейронов, иннервирующих сердце и бронхи. Эффекторные нейроны этого пути располагаются в звездчатом ганглии или в узлах пограничного симпатического ствола. Стимуляция этих волокон и клеток вызывает учащение и усиление сердечных сокращений и расширение бронхов.
На уровне всех грудных, а также верхних поясничных сегментов спинного мозга, т. е. на всем протяжении симпатического ядра, расположены нейроны, иннервирующие сосуды и потовые железы. Характерной чертой этих скоплений нейронов является топография клеточных тел и определяемая ею зона иннервации. Поражение клеточных скоплений отдельных сегментов, как и их разрушение, сопровождается исчезновением потоотделения.
Крестцовые отделы спинного мозга занимают парасимпатические нейроны. Их совокупности образуют ряд центров рефлексов мочеиспускания, дефекации, эрекции и т. д. Поражение этих центров ведет к выпадению названных функций.
Стволовые центры. Располагающиеся в продолговатом мозге, мосте и среднем мозге скопления парасимпатических нейронов образуют центры, в которых осуществляется замыкание рефлексов сосания, жевания, глотания, чиханья, кашля, рвоты, слюноотделения, слезотечения, торможения сердечной деятельности, секреции желудочных желез и т. д. Эти влияния передаются исполнительным структурам по волокнам блуждающего, языкоглоточного, лицевого и глазодвигательного нервов.
Расположение этих центров непостоянно, составляющие их группы нейронов небольшие и не различаются морфологически. Кроме того, клетки, управляющие какой-либо определенной функцией, располагаются не всегда вместе и рядом. Следовательно, понятие «центр», означающее функционально связанные совокупности нейронов, расположенные в одном или нескольких структурах ЦНС и обеспечивающие существование целостной реакции организма или регуляцию определенной функции, может использоваться в этом случае с определенными допущениями. В продолговатом мозге в ядрах блуждающего нерва замыкаются рефлексы с аортальной и синокаротидной рефлексогенных зон, рефлекс снижения частоты сердечных сокращений при раздражении интероцепторов брюшной полости (рефлекс Гольца), глазосердечный рефлекс (рефлекс Ашнера).
Часто рефлекторные реакции сердца проявляются сопряженно с изменением сосудистого тонуса, что определяется наличием связей между нейронами, которые регулируют сердечную деятельность и сосудистый тонус. Волокна блуждающего нерва несут импульсы, управляющие деятельностью системы дыхания, пищеварения. Центры, регулирующие работу слюнных желез, осуществляют свое влияние по нервным волокнам, следующим в составе языкоглоточного и лицевого нервов, а центры зрачкового рефлекса и рефлекса аккомодации глаза располагаются в среднем мозге, передних буграх четверохолмия. Импульсы к слезной железе следуют по веточкам лицевого нерва.
Сосудодвигательный центр — морфофункциональное образование продолговатого мозга, играющее ведущую роль в поддержании тонуса сосудов и регуляции кровяного давления. Он координирует и деятельность спинномозгового симпатического центра, посылающего сосудосуживающие импульсы к сосудистой стенке. Тонус сосудодвигательного центра и, следовательно, уровень общего артериального давления регулируется импульсами, возникающими в сосудистых рефлексогенных зонах. Сосудодвигательный центр входит в состав ретикулярной формации продолговатого мозга и поэтому получает многочисленные коллатеральные возбуждения от всех специфических проводящих путей, что постоянно поддерживает его в состоянии тонического возбуждения.
В ответах всего организма сосудодвигательный центр выступает в качестве исполнительного органа, через который в значительной мере реализуются супрабульбарные влияния на гемодинамику. Влияние самого центра осуществляется через спинной мозг, периферические симпатические образования, блуждающие нервы и обусловливает преимущественно системные изменения гемодинамики. Считают, что в любых случаях нейрогенная гипертензия обусловлена стойким повышением возбудимости бульварных сосудосуживающих структур.
Рефлекторные процессы в ядерных образованиях спинного, продолговатого, среднего мозга и моста находятся под постоянным влиянием гипоталамуса.
Гипоталамические центры. Гипоталамусу принадлежит ведущая роль в осуществлении многих функций целого организма и прежде всего постоянства внутренней среды. В нем осуществляется интеграция и приспособление различных висцеральных систем к целостной деятельности организма.
В гипоталамусе принято различать три нерезко ограниченные области скопления ядер: переднюю, среднюю и заднюю. Гипоталамус обладает хорошо развитой сложной системой афферентных и эфферентных путей, а также тесно связан со структурами головного мозга — таламусом, лимбической системой, ретикулярной формацией ствола мозга. Особое значение имеют обширные сосудистые и нервные связи с гипофизом, в результате чего осуществляется интегрирование нервной и гуморальной регуляции висцеральных функций. Гипоталамусом она осуществляется двумя путями: парааденогипофизарным (минуя аденогипофиз) и трансаденогипофизарным (через аденогипофиз). Такие многочисленные связи гипоталамуса с другими образованиями мозга способствуют генерализации возбуждения, возникающего в его нейронах. Возбуждение в первую очередь распространяется на лимбические структуры мозга и через ядра таламуса на передние отделы коры большого мозга. Результаты раздражения структур гипоталамуса определяются его контактами с ретикулярной формацией, симпатическими и парасимпатическими центрами, а также усилением секреции гормонов гипофиза, действующих непосредственно или опосредованно через другие эндокринные железы. Следовательно, при стимуляции гипоталамуса возникают сложные реакции, в которых нервный компонент дополняется гормональным. Регуляция гипоталамо-гипофизарной системой висцеральных функций осуществляется по принципу обратной связи.
Активация гипоталамических ядер зависит не только от поступления к ним возбуждающих влияний из других структур нервной системы, но и избирательной чувствительности их клеток к содержанию тех или иных веществ в крови, изменению температуры крови. Например, гипоталамические нейроны чувствительны к малейшим отклонениям рН крови, напряжению О2, СО2, содержанию ионов, особенно калия и натрия. В супраоптическом ядре имеются клетки, избирательно чувствительные к изменению осмотического давления крови, в вентромедиальном ядре — к содержанию глюкозы, в переднем гипоталамусе — половых органов. Таким образом, клетки гипоталамуса наряду с другими свойствами выполняют рецепторные функции, воспринимая нарушения гомеостаза. Они обладают способностью трансформировать гуморальные изменения внутренней среды в нервный процесс. Кроме того, они могут избирательно активироваться нервными импульсами из соответствующих органов.
При стимуляции гипоталамуса возникает комплекс сложных реакций, в которых нервный компонент дополняется гормональным. Так, раздражение ядер задней группы характеризуется эффектами, аналогичными раздражению симпатической части автономной нервной системы — расширяются зрачки и глазная щель, возрастает частота сердечных сокращений, повышается кровяное давление, тормозится двигательная активность пищеварительного тракта, в крови возрастает концентрация адреналина и норадреналина. Разрушение этой области приводит к гипергликемии, ожирению, нарушению терморегуляции.
Раздражение ядер передней группы сопровождается реакциями, подобными в определенной мере раздражению парасимпатической части автономной нервной системы, — сужением зрачков и глазных щелей, урежением частоты сердечных сокращений, снижением артериального давления, усилением двигательной активности желудочно-кишечного тракта. Ядра этой группы участвуют в механизме терморегуляции.
Функция ядер средних групп состоит преимущественно в регуляции метаболизма. Разрушение, например, вентромедиальных ядер сопровождается повышением потребления пищи (гиперфагия) и ожирением, двустороннее разрушение латеральных ядер, напротив, приводит к полному отказу от пищи. Эти показатели явились основанием расценивать вентромедиальные ядра как структуры, связанные с насыщением, а латеральные ядра — с голодом. Наибольшую потребность в воде (полидипсия) регистрировали при раздражении паравентрикулярного ядра гипоталамуса. При хронической стимуляции ядер этой группы у животных возникают атеросклеротические изменения сосудов.
Стимуляция ядер гипоталамуса независимо от того, к какой топографической группе они относятся, непременно сопровождается сложными гормональными реакциями: увеличиваются выделение тропных гормонов передней доли гипофиза, секреция задней доли. В ответных реакциях организма при раздражении разных областей гипоталамуса участвуют практически все висцеральные органы, изменяются поведенческие реакции, эмоциональная деятельность целого организма. Используя нейротропные препараты, можно избирательно блокировать гипоталамические механизмы формирования состояния голода, жажды, аппетита, страха, половых и агрессивно-оборонительных реакций.
Гипоталамус связан прямыми нервными путями и через ретикулярную формацию ствола мозга с подкорковыми ядрами, мозжечком, корой больших полушарий. Его деятельность постоянно контролируется высшими центрами ЦНС. Гипоталамус занимает ведущее место в регуляции функций организма и прежде всего постоянства внутренней среды. Под его контролем находится функция автономной нервной системы и эндокринных желез.
Лимбическая система. Связь функций лимбической системы с работой внутренних органов послужила основанием для обозначения совокупности ее структур термином «висцеральный мозг».
Лимбическая система обеспечивает взаимодействие экстероцептивных (обонятельных, слуховых и др.) и интероцептивных воздействий. Она регулирует висцерально-гормональные функции, направленные на обеспечение различных форм деятельности, таких, как пищевое, сексуальное, оборонительное поведение, регулирует системы, обеспечивающие сон и бодрствование, внимание, эмоциональную сферу, процессы памяти, осуществляя, таким образом, соматовисцеральную интеграцию (подробнее см. раздел 4.2.3).
Мозжечок. Наряду с регуляцией двигательной соматической сферы мозжечок контролирует течение висцеральных процессов. При его раздражении могут быть воспроизведены практически все реакции, возникающие при возбуждении симпатической нервной системы — расширение зрачка, сужение сосудов, сокращение волосяных мышц, учащение сердечного ритма. После удаления мозжечка возникает угнетение периодической моторной деятельности пищеварительного тракта, секреторной функции кишечных желез и т. д. Это указывает на то, что мозжечок благодаря наличию активирующего и тормозного механизмов может оказывать в целом организме стабилизирующее влияние на деятельность висцеральных органов посредством корригирования висцеральных рефлексов.
Ретикулярная формация. Основной ролью ее нисходящей части по отношению к деятельности автономной нервной системы является повышение активности нервных центров, связанных с висцеральными функциями. Ретикулярная формация оказывает на них тонизирующее влияние, обеспечивая высокий уровень их активности. Проводником этих влияний на периферию является симпатическая часть автономной нервной системы. В поддержании активности ретикулярных механизмов значительную роль играют гуморальные раздражения, по отношению к которым она обладает высокой чувствительностью. Сами же функциональные влияния ретикулярной формации сказываются и на эндокринном компоненте регуляции поведения висцеральных систем. Действительно при стимуляции ретикулярной формации среднего и промежуточного мозга усиливается выброс гипофизарных гормонов, а при нарушении этих ретикулярных образований возникают эндокринные расстройства.
Кора большого мозга. Еще в середине прошлого столетия благодаря исследованиям В. Я. Данилевского, В. М. Бехтерева, Н. А. Миславского стало известно, что раздражение или выключение отдельных участков коры большого мозга влечет за собой изменение состояний внутренних органов. При этом могли быть зарегистрированы противоположные по направленности изменения висцеральных функций типа повышения или снижения кровяного давления, усиления или ослабления моторной активности органов пищеварения.
У человека раздражение коры кзади от центральной (роландовой) борозды и вблизи латеральной (сильвиевой) борозды вызывает ощущение тошноты, рвоты, возникают позывы на дефекацию. Раздражение точек в теменных и других долях сопровождается изменением сердечной деятельности, артериального давления, дыхательного ритма, слюноотделения, желудочной и кишечной моторики.
Особое значение в регуляции функций в настоящее время придается лобным долям коры большого мозга, поскольку при их стимуляции можно зарегистрировать изменение практически всех висцеральных процессов. Именно из-за этого передние отделы больших полушарий считаются высшими центрами автономной иннервации. Однако наряду с этим существует определенная специализация некоторых полей коры. Так, в ее двигательных областях находится представительство тех висцеральных органов, деятельность которых связана со скелетно-мышечной активностью. Посредством такой организации достигается необходимая для нормальной жизнедеятельности интеграция соматических и висцеральных процессов.
Исследования В. Н. Черниговским интероцепции и представительства в коре большого мозга висцеральных систем показали, что в определенных условиях информация о работе внутренних органов может достигать высших отделов ЦНС. Тем самым было экспериментально подтверждено выдвинутое И. П. Павловым понятие о корковом представительстве интероцептивного анализатора.
Известно, что в определенных условиях у человека гипнотическим внушением можно вызвать изменение сердечного ритма, вазоконстрикцию и вазодилатацию, усиление поте- и мочеотделения, изменение метаболизма.
К. М. Быков обосновал возможность образования висцеральных условных рефлексов влиянием коры большого мозга на деятельность внутренних органов. Это легло в основу концепции существования кортико-висцеральных отношений. Сейчас они рассматриваются не более как способы модуляции корой деятельности подкорковых структур, имеющих непосредственное отношение к регуляции внутренней среды организма.
Физиология человека, под редакцией В. М. Покровского, Г. Ф. Коротько
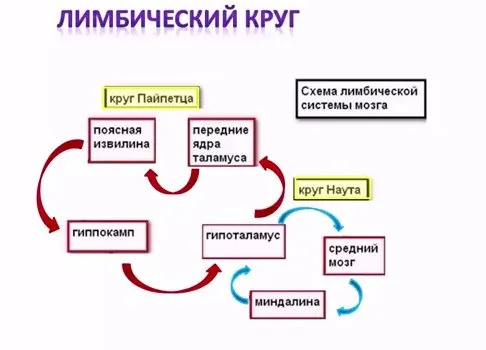
Сайт Светланы Анатольевны Коппел-Ковтун

Комментарии
Лимбическая система - эмоциональный мозг (чувствуем в области сердца и груди), а есть ещё интеллектуальный и брюшной (мозг дела и тела). Мозг - это выживание, и у каждого отдела мозга свои инструменты. Брюшной (у рептилий): инструменты выживания - инстинкты (5 инстинктов поддерживающих жизнь и 5 инстинктов, которые включаются в момент угрозы выживанию).
Лимбическая (мозг млекопитающих) - инструментами выживания являются эмоции и чувства. Любовь, радость - у рептилий этого нету, они не переживают эмоцию, у них нет привязанностей. Они не заступаются друг за друга. У млекопитающих появляется лимбическая система, которая создала другой инструмент - эмоции и чувства. Млекопитающие дружат, собираются в стаи и стада, защищают потомство.
Инструментом неокортекса (новой коры) является создание ассоциативных связок. Неокортекс позволяет связывать вещи и явления друг с другом.
- Ответить
Ссылка на комментарийЗдравствуйте, Светлана!
Наткнулась на данную статью и приступила к ее планомерному изучению и обдумыванию. У меня вопрос относительно рептильного мозга и Вашего комментария. Верно ли я понимаю, что рептильный мозг и брюшной мозг вещи разные и между ними имеется взаимосвязь, являющаяся древней эволюционной механикой функционирования организма? Или я неверно трактую эти понятия?
Буду благодарна за ответ.
- Ответить
Ссылка на комментарийЯ в этом не спец, сама пыталась разобраться, но насколько я поняла, как раз рептильный и есть брюшной. Мне это интересно в связи с грехопадением. Ева в раю говорит со Змием. Как он туда попал? Я думаю, что наш кишечник - это по сути змея, как по Сеченову мышца - это животное. Отсюда можно предполагать, что контакт Евы с этим древним Змием надо искать не в той плоскости, что ищут обычно.
- Ответить
Ссылка на комментарийОставить комментарий